Evaluation of Active Brown Adipose Tissue by the Use of Hyperpolarized [1-13C]Pyruvate MRI in Mice

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
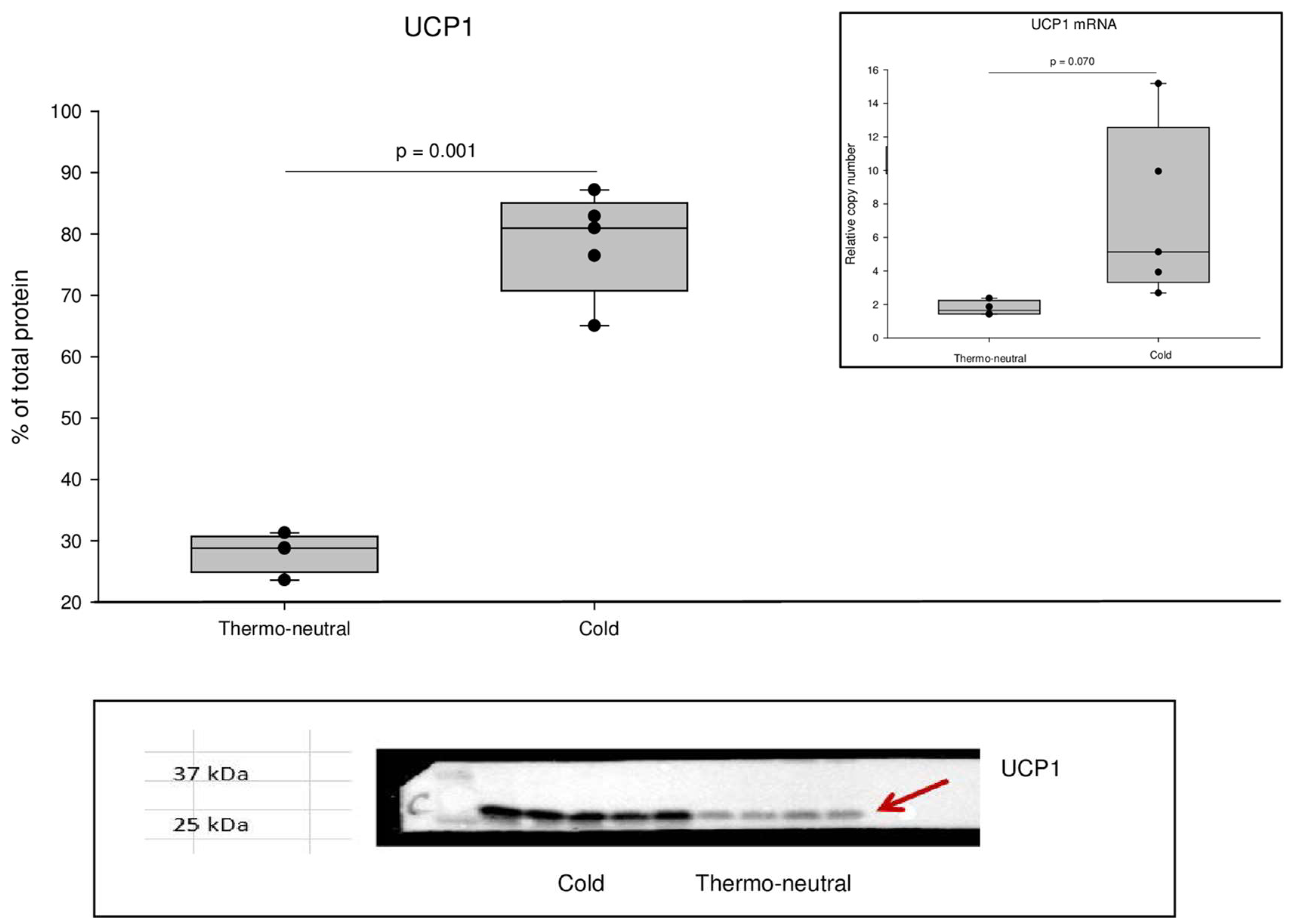
2.1. UCP1 Levels in iBAT
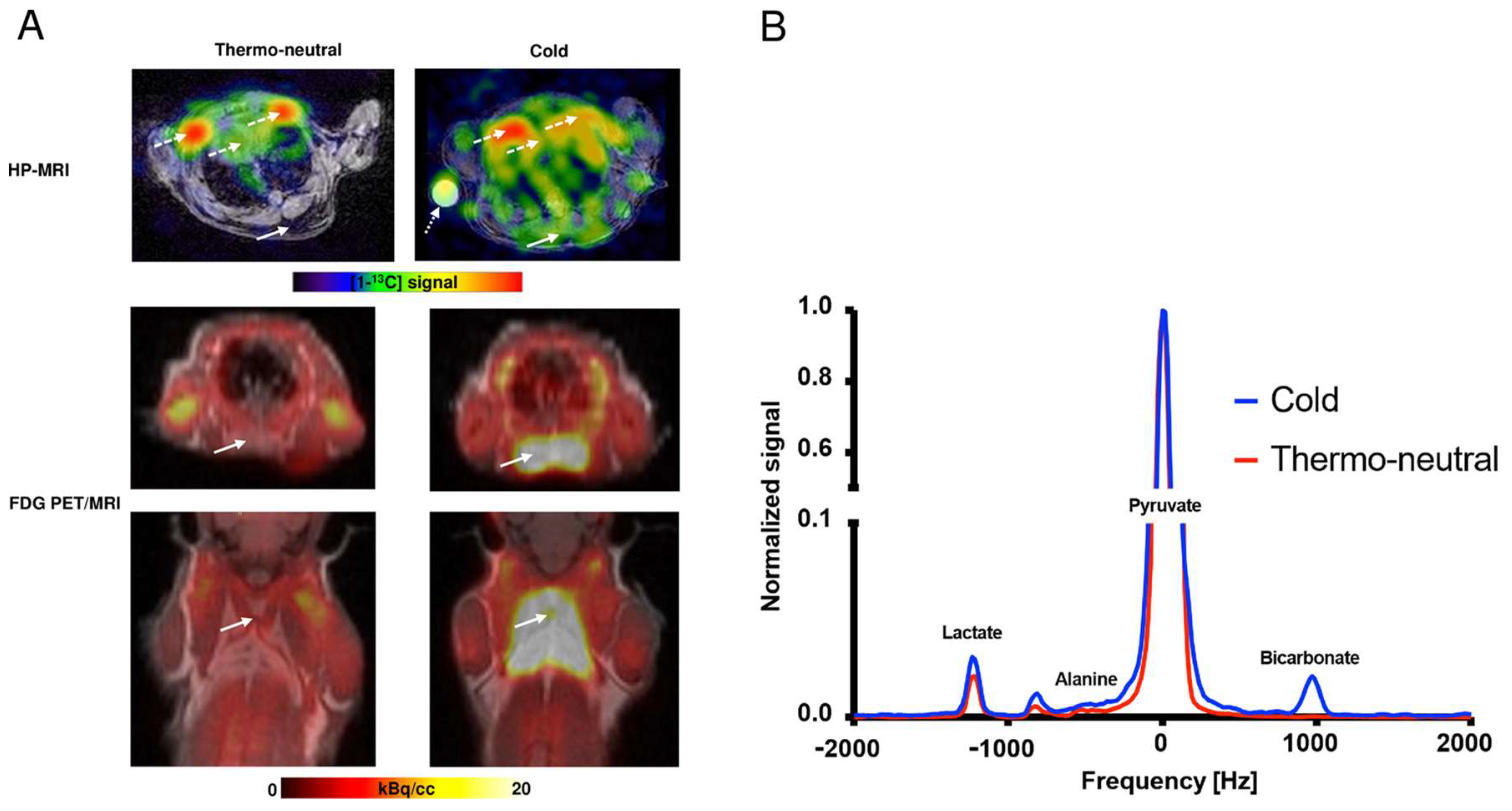
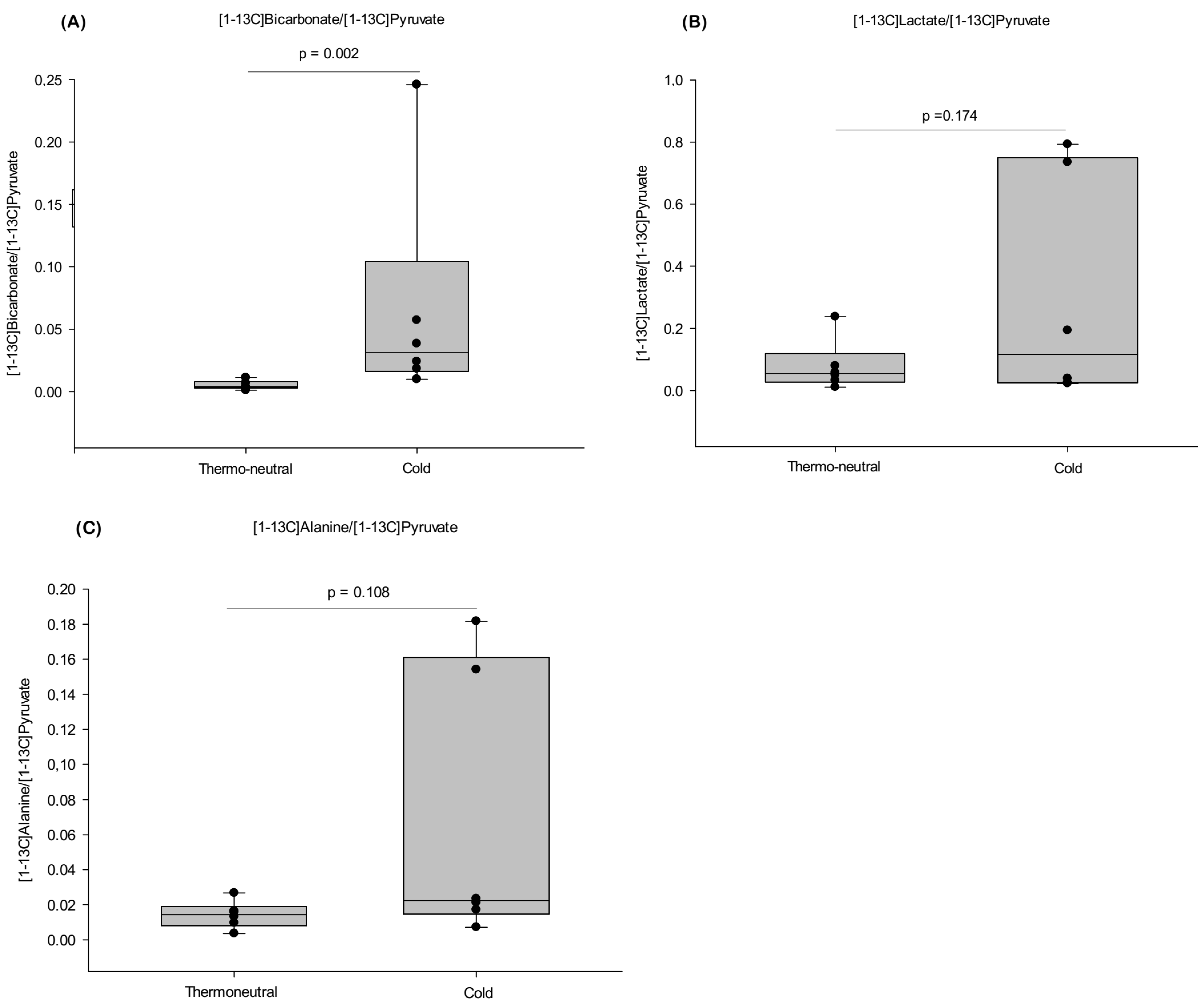
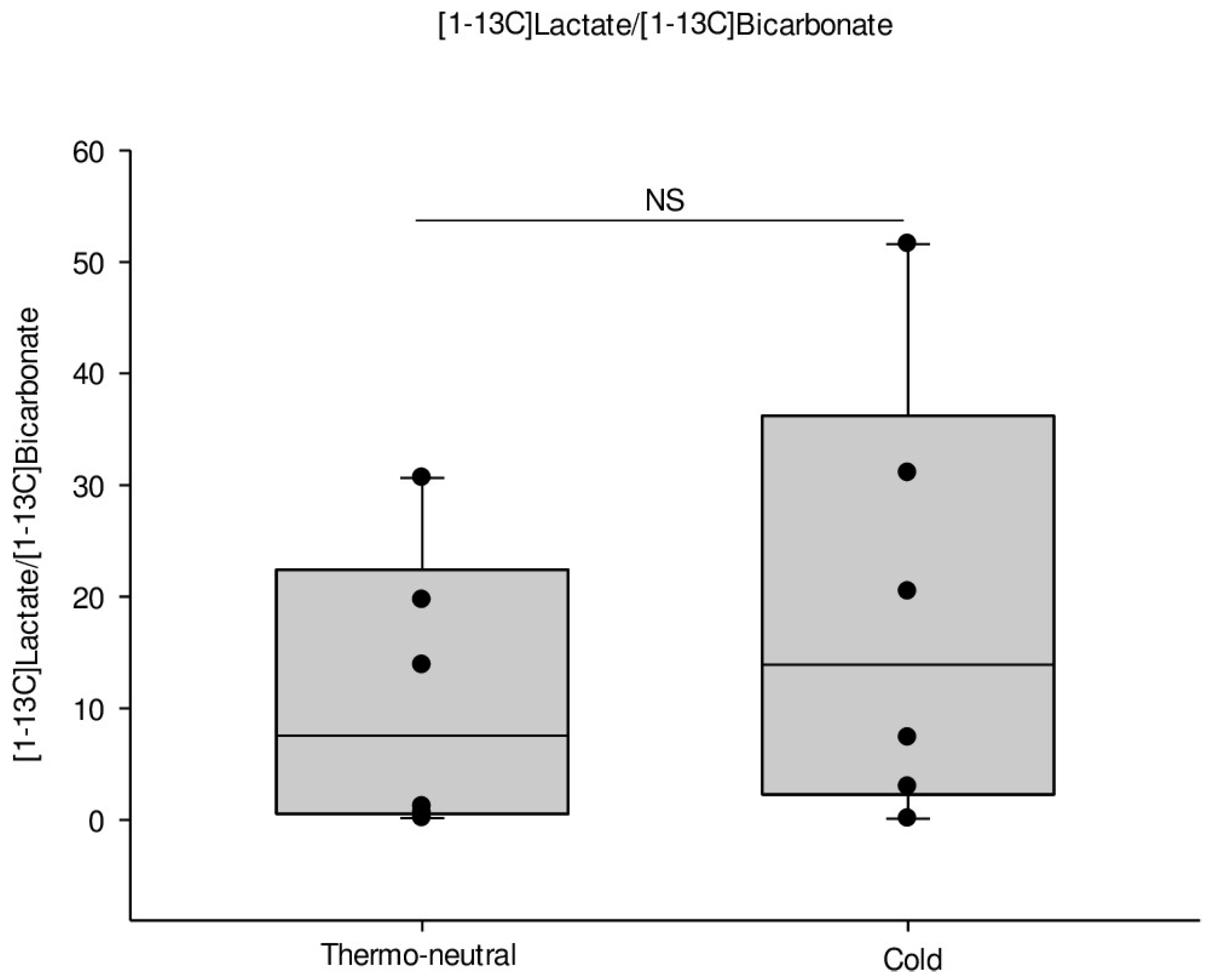
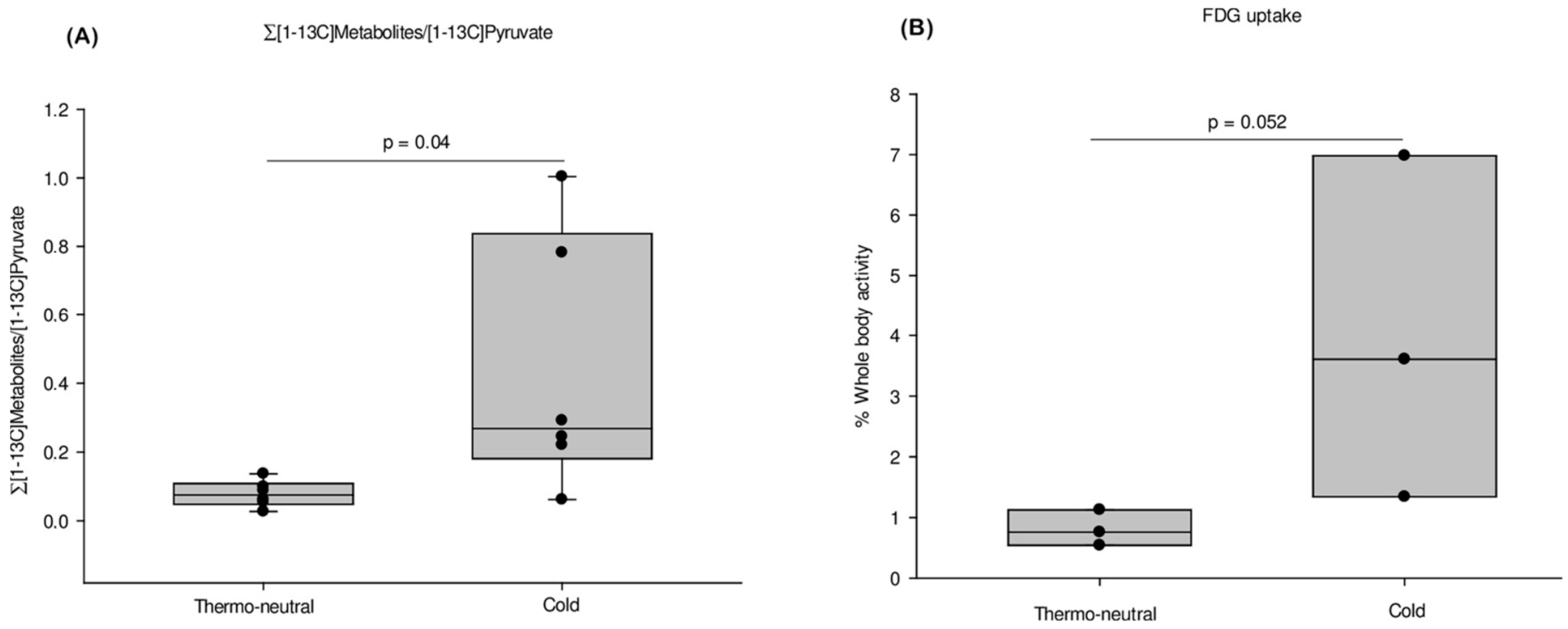
2.2. HP-MRI Determination of iBAT Activity
2.3. FDG PET/MRI Determination of iBAT Activity
3. Discussion
4. Materials and Methods
4.1. Animal Handling
4.2. Protocol for Temperature Acclimatization
4.3. HP [1-13C]pyruvate-MRI
4.4. FDG PET/MRI
4.5. Gene Expression and Protein Determination
4.6. Statistical Method
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of interest
References
- Norris, S.L.; Zhang, X.; Avenell, A.; Gregg, E.; Bowman, B.; Serdula, M.; Brown, T.J.; Schmid, C.H.; Lau, J. Long-Term Effectiveness of Lifestyle and Behavioral Weight Loss Interventions in Adults with Type 2 Diabetes: A Meta-Analysis. Am. J. Med. 2004, 117, 762–774. [Google Scholar] [CrossRef] [PubMed]
- Franz, M.J.; Boucher, J.L.; Rutten-Ramos, S.; VanWormer, J.J. Lifestyle Weight-Loss Intervention Outcomes in Overweight and Obese Adults with Type 2 Diabetes: A Systematic Review and Meta-Analysis of Randomized Clinical Trials. J. Acad. Nutr. Diet. 2015, 115, 1447–1463. [Google Scholar] [CrossRef] [PubMed]
- Koksharova, E.; Ustyuzhanin, D.; Philippov, Y.; Mayorov, A.; Shestakova, M.; Shariya, M.; Ternovoy, S.; Dedov, I. The Relationship between Brown Adipose Tissue Content in Supraclavicular Fat Depots and Insulin Sensitivity in Patients with Type 2 Diabetes Mellitus and Prediabetes. Diabetes Technol. Ther. 2017, 19, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Romu, T.; Vavruch, C.; Dahlqvist-Leinhard, O.; Tallberg, J.; Dahlstrom, N.; Persson, A.; Heglind, M.; Lidell, M.E.; Enerback, S.; Borga, M.; et al. A Randomized Trial of Cold-Exposure on Energy Expenditure and Supraclavicular Brown Adipose Tissue Volume in Humans. Metabolism 2016, 65, 926–934. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.W.; Foster, D.O. Uptake of Glucose and Release of Fatty Acids and Glycerol by Rat Brown Adipose Tissue in Vivo. Can. J. Physiol. Pharmacol. 1986, 64, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Fukano, K.; Okamatsu-Ogura, Y.; Tsubota, A.; Nio-Kobayashi, J.; Kimura, K. Cold Exposure Induces Proliferation of Mature Brown Adipocyte in a β3-Adrenergic Receptor-Mediated Pathway. PLoS ONE 2016, 11, e0166579. [Google Scholar] [CrossRef] [PubMed]
- Brychta, R.J.; Chen, K.Y. Cold-Induced Thermogenesis in Humans. Eur. J. Clin. Nutr. 2017, 71, 345. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P. Origins and Early Development of the Concept that Brown Adipose Tissue Thermogenesis is Linked to Energy Balance and Obesity. Biochimie 2017, 134, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Cypess, A.M.; Lehman, S.; Williams, G.; Tal, I.; Rodman, D.; Goldfine, A.B.; Kuo, F.C.; Palmer, E.L.; Tseng, Y.H.; Doria, A.; et al. Identification and Importance of Brown Adipose Tissue in Adult Humans. N. Engl. J. Med. 2009, 360, 1509–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virtanen, K.A.; Lidell, M.E.; Orava, J.; Heglind, M.; Westergren, R.; Niemi, T.; Taittonen, M.; Laine, J.; Savisto, N.J.; Enerback, S.; et al. Functional Brown Adipose Tissue in Healthy Adults. N. Engl. J. Med. 2009, 360, 1518–1525. [Google Scholar] [CrossRef] [PubMed]
- Van Marken Lichtenbelt, W.D.; Vanhommerig, J.W.; Smulders, N.M.; Drossaerts, J.M.; Kemerink, G.J.; Bouvy, N.D.; Schrauwen, P.; Teule, G.J. Cold-Activated Brown Adipose Tissue in Healthy Men. N. Engl. J. Med. 2009, 360, 1500–1508. [Google Scholar] [CrossRef] [PubMed]
- Blondin, D.P.; Daoud, A.; Taylor, T.; Tingelstad, H.C.; Bezaire, V.; Richard, D.; Carpentier, A.C.; Taylor, A.W.; Harper, M.E.; Aguer, C.; et al. Four-Week Cold Acclimation in Adult Humans Shifts Uncoupling Thermogenesis from Skeletal Muscles to Brown Adipose Tissue. J. Physiol. 2017, 595, 2099–2113. [Google Scholar] [CrossRef] [PubMed]
- Murholm, M.; Isidor, M.S.; Basse, A.L.; Winther, S.; Sorensen, C.; Skovgaard-Petersen, J.; Nielsen, M.M.; Hansen, A.S.; Quistorff, B.; Hansen, J.B. Retinoic Acid has Different Effects on UCP1 Expression in Mouse and Human Adipocytes. BMC Cell Biol. 2013, 14, 41. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.I.; Cypess, A.M.; Sass, C.A.; Brownell, A.L.; Jokivarsi, K.T.; Kahn, C.R.; Kwong, K.K. Anatomical and Functional Assessment of Brown Adipose Tissue by Magnetic Resonance Imaging. Obesity 2012, 20, 1519–1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virtanen, K.A. The Rediscovery of BAT in Adult Humans using Imaging. Best Pract. Res. Clin. Endocrinol. Metab. 2016, 30, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.Y.; Cypess, A.M.; Laughlin, M.R.; Haft, C.R.; Hu, H.H.; Bredella, M.A.; Enerback, S.; Kinahan, P.E.; Lichtenbelt, W.; Lin, F.I.; et al. Brown Adipose Reporting Criteria in Imaging STudies (BARCIST 1.0): Recommendations for Standardized FDG-PET/CT Experiments in Humans. Cell Metab. 2016, 24, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Gatidis, S.; Schmidt, H.; Pfannenberg, C.A.; Nikolaou, K.; Schick, F.; Schwenzer, N.F. Is it Possible to Detect Activated Brown Adipose Tissue in Humans Using Single-Time-Point Infrared Thermography Under Thermoneutral Conditions? Impact of BMI and Subcutaneous Adipose Tissue Thickness. PLoS ONE 2016, 11, e0151152. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.W.; Ootsuka, Y.; Romanovsky, A.A. Body Temperature Measurements for Metabolic Phenotyping in Mice. Front. Physiol. 2017, 8, 520. [Google Scholar] [CrossRef] [PubMed]
- Malloy, C.R.; Sherry, A.D. Biochemical Specificity in Human Cardiac Imaging by 13C Magnetic Resonance Imaging. Circ. Res. 2016, 119, 1146–1148. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, C.; Lycke, S.; Palm, F.; Ostergaard, J.A.; Bibby, B.M.; Norregaard, R.; Flyvbjerg, A.; Pedersen, M.; Ardenkjaer-Larsen, J.H. High Altitude may Alter Oxygen Availability and Renal Metabolism in Diabetics as Measured by Hyperpolarized [1-13C]Pyruvate Magnetic Resonance Imaging. Kidney Int. 2014, 86, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Hurd, R.E.; Yen, Y.F.; Chen, A.; Ardenkjaer-Larsen, J.H. Hyperpolarized 13C Metabolic Imaging Using Dissolution Dynamic Nuclear Polarization. J. Magn. Reson. Imaging 2012, 36, 1314–1328. [Google Scholar] [CrossRef] [PubMed]
- Lau, A.Z.; Chen, A.P.; Gu, Y.; Ladouceur-Wodzak, M.; Nayak, K.S.; Cunningham, C.H. Noninvasive Identification and Assessment of Functional Brown Adipose Tissue in Rodents using Hyperpolarized 13C Imaging. Int. J. Obes. 2014, 38, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, S.; Kadlecek, S.; Pourfathi, M.; Xin, Y.; Mannherz, W.; Hamedani, H.; Drachman, N.; Ruppert, K.; Clapp, J.; Rizi, R. The Use of Hyperpolarized Carbon-13 Magnetic Resonance for Molecular Imaging. Adv. Drug Deliv. Rev. 2017, 113, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Nelson, S.J.; Kurhanewicz, J.; Vigneron, D.B.; Larson, P.E.; Harzstark, A.L.; Ferrone, M.; van Criekinge, M.; Chang, J.W.; Bok, R.; Park, I.; et al. Metabolic Imaging of Patients with Prostate Cancer Using Hyperpolarized [1-13C]Pyruvate. Sci. Transl. Med. 2013, 5, 198ra108. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, C. Hyperpolarized Renal Magnetic Resonance Imaging: Potential and Pitfalls. Front. Physiol. 2016, 7, 72. [Google Scholar] [CrossRef] [PubMed]
- Ball, D.R.; Cruickshank, R.; Carr, C.A.; Stuckey, D.J.; Lee, P.; Clarke, K.; Tyler, D.J. Metabolic Imaging of Acute and Chronic Infarction in the Perfused Rat Heart Using Hyperpolarised [1-13C]Pyruvate. NMR Biomed. 2013, 26, 1441–1450. [Google Scholar] [CrossRef] [PubMed]
- Townsend, K.L.; Tseng, Y.H. Brown Fat Fuel Utilization and Thermogenesis. Trends Endocrinol. Metab. 2014, 25, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, P.M.; Laustsen, C.; Bertelsen, L.B.; Qi, H.; Mikkelsen, E.; Kristensen, M.L.; Norregaard, R.; Stodkilde-Jorgensen, H. In Situ Lactate Dehydrogenase Activity: A Novel Renal Cortical Imaging Biomarker of Tubular Injury? Am. J. Physiol. Renal Physiol. 2017, 312, F465–F473. [Google Scholar] [CrossRef] [PubMed]
- Ohlson, K.B.; Lindahl, S.G.; Cannon, B.; Nedergaard, J. Thermogenesis Inhibition in Brown Adipocytes is a Specific Property of Volatile Anesthetics. Anesthesiology 2003, 98, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Albanese, C.T.; Nour, B.M.; Rowe, M.I. Anesthesia Blocks Nonshivering Thermogenesis in the Neonatal Rabbit. J. Pediatr. Surg. 1994, 29, 983–986. [Google Scholar] [CrossRef]
- McCallister, A.; Zhang, L.; Burant, A.; Katz, L.; Branca, R.T. A Pilot Study on the Correlation between Fat Fraction Values and Glucose Uptake Values in Supraclavicular Fat by Simultaneous PET/MRI. Magn. Reson. Med. 2017, 78, 1922–1932. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, P.M.; Szocska Hansen, E.S.; Norlinger, T.S.; Norregaard, R.; Bonde Bertelsen, L.; Stodkilde Jorgensen, H.; Laustsen, C. Renal Ischemia and Reperfusion Assessment with Three-Dimensional Hyperpolarized 13C,15N2-Urea. Magn. Reson. Med. 2016, 76, 1524–1530. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.H.; Lau, J.Y.; Chen, A.P.; Geraghty, B.J.; Perks, W.J.; Roifman, I.; Wright, G.A.; Connelly, K.A. Hyperpolarized 13C Metabolic MRI of the Human Heart: Initial Experience. Circ. Res. 2016, 119, 1177–1182. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, R.; Vigneron, D.B.; Kurhanewicz, J. Hyperpolarized [1-13C]Pyruvate Magnetic Resonance Imaging Detects an Early Metabolic Response to Androgen Ablation Therapy in Prostate Cancer. Eur. Urol. 2017, 72, 1028–1029. [Google Scholar] [CrossRef] [PubMed]
- Balzan, R.; Fernandes, L.; Pidial, L.; Comment, A.; Tavitian, B.; Vasos, P.R. Pyruvate Cellular Uptake and Enzymatic Conversion Probed by Dissolution DNP-NMR: The Impact of Overexpressed Membrane Transporters. Magn. Reson. Chem. 2017, 55, 579–583. [Google Scholar] [CrossRef] [PubMed]
- Keshari, K.R.; Sriram, R.; van Criekinge, M.; Wilson, D.M.; Wang, Z.J.; Vigneron, D.B.; Peehl, D.M.; Kurhanewicz, J. Metabolic Reprogramming and Validation of Hyperpolarized 13C Lactate as a Prostate Cancer Biomarker using a Human Prostate Tissue Slice Culture Bioreactor. Prostate 2013, 73, 1171–1181. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, C.; Nielsen, P.M.; Norlinger, T.S.; Qi, H.; Pedersen, U.K.; Bertelsen, L.B.; Ostergaard, J.A.; Flyvbjerg, A.; Ardenkjaer-Larsen, J.H.; Palm, F.; et al. Antioxidant Treatment Attenuates Lactate Production in Diabetic Nephropathy. Am. J. Physiol. Renal Physiol. 2017, 312, F192–F199. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, C.; Ostergaard, J.A.; Lauritzen, M.H.; Norregaard, R.; Bowen, S.; Sogaard, L.V.; Flyvbjerg, A.; Pedersen, M.; Ardenkjaer-Larsen, J.H. Assessment of Early Diabetic Renal Changes with Hyperpolarized [1-13C]Pyruvate. Diabetes Metab. Res. Rev. 2013, 29, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Laustsen, C.; Dumez, J.N.; Kettunen, M.I.; Serrao, E.M.; Marco-Rius, I.; Brindle, K.M.; Ardenkjaer-Larsen, J.H.; Frydman, L. In Vivo Single-Shot 13C Spectroscopic Imaging of Hyperpolarized Metabolites by Spatiotemporal Encoding. J. Magn. Reson. 2014, 240, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Hansen, E.S.; Kim, S.; Miller, J.J.; Geferath, M.; Morrell, G.; Laustsen, C. Fast Pade Transform Accelerated CSI for Hyperpolarized MRS. Tomography 2016, 2, 117–124. [Google Scholar] [PubMed]
- Vinding, M.S.; Laustsen, C.; Maximov, I.I.; Sogaard, L.V.; Ardenkjaer-Larsen, J.H.; Nielsen, N.C. Dynamic Nuclear Polarization and Optimal Control Spatial-Selective 13C MRI and MRS. J. Magn. Reson. 2013, 227, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.P.; Cunningham, C.H. Single Voxel Localization for Dynamic Hyperpolarized 13C MR Spectroscopy. J. Magn. Reson. 2015, 258, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Jung, K.H.; Lee, J.H.; Quach, C.H.; Moon, S.H.; Cho, Y.S.; Lee, K.H. 18F-FDG PET/CT Monitoring of β3 Agonist-Stimulated Brown Adipocyte Recruitment in White Adipose Tissue. J. Nucl. Med. 2015, 56, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Hankir, M.K.; Kranz, M.; Keipert, S.; Weiner, J.; Andreasen, S.G.; Kern, M.; Patt, M.; Kloting, N.; Heiker, J.T.; Hesse, S.; et al. Dissociation between Brown Adipose Tissue 18F-FDG Uptake and Thermogenesis in Uncoupling Protein 1 Deficient Mice. J. Nucl. Med. 2017, 58, 1100–1103. [Google Scholar] [CrossRef] [PubMed]
- Breining, P.; Pedersen, S.B.; Pikelis, A.; Rolighed, L.; Ordell Sundelin, E.I.; Jessen, N.; Richelsen, B. High Expression of Organic Cation Transporter 3 in Human BAT-Like Adipocytes. Implications for Extraneuronal Norepinephrine Uptake. Mol. Cell. Endocrinol. 2017, 443, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Arvidsson, S.; Kwasniewski, M.; Riano-Pachon, D.M.; Mueller-Roeber, B. QuantPrime—A Flexible Tool for Reliable High-Throughput Primer Design for Quantitative PCR. BMC Bioinformatics 2008, 9, 465. [Google Scholar] [CrossRef] [PubMed]
- Sondergaard, E.; Gormsen, L.C.; Christensen, M.H.; Pedersen, S.B.; Christiansen, P.; Nielsen, S.; Poulsen, P.L.; Jessen, N. Chronic Adrenergic Stimulation Induces Brown Adipose Tissue Differentiation in Visceral Adipose Tissue. Diabet. Med. 2015, 32, e4–e8. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riis-Vestergaard, M.J.; Breining, P.; Pedersen, S.B.; Laustsen, C.; Stødkilde-Jørgensen, H.; Borghammer, P.; Jessen, N.; Richelsen, B. Evaluation of Active Brown Adipose Tissue by the Use of Hyperpolarized [1-13C]Pyruvate MRI in Mice. Int. J. Mol. Sci. 2018, 19, 2597. https://doi.org/10.3390/ijms19092597
Riis-Vestergaard MJ, Breining P, Pedersen SB, Laustsen C, Stødkilde-Jørgensen H, Borghammer P, Jessen N, Richelsen B. Evaluation of Active Brown Adipose Tissue by the Use of Hyperpolarized [1-13C]Pyruvate MRI in Mice. International Journal of Molecular Sciences. 2018; 19(9):2597. https://doi.org/10.3390/ijms19092597
Chicago/Turabian StyleRiis-Vestergaard, Mette Ji, Peter Breining, Steen Bønløkke Pedersen, Christoffer Laustsen, Hans Stødkilde-Jørgensen, Per Borghammer, Niels Jessen, and Bjørn Richelsen. 2018. "Evaluation of Active Brown Adipose Tissue by the Use of Hyperpolarized [1-13C]Pyruvate MRI in Mice" International Journal of Molecular Sciences 19, no. 9: 2597. https://doi.org/10.3390/ijms19092597