Neglected Functions of TFCP2/TFCP2L1/UBP1 Transcription Factors May Offer Valuable Insights into Their Mechanisms of Action


{kind=link}
{kind=link}
Abstract
:1. Introduction
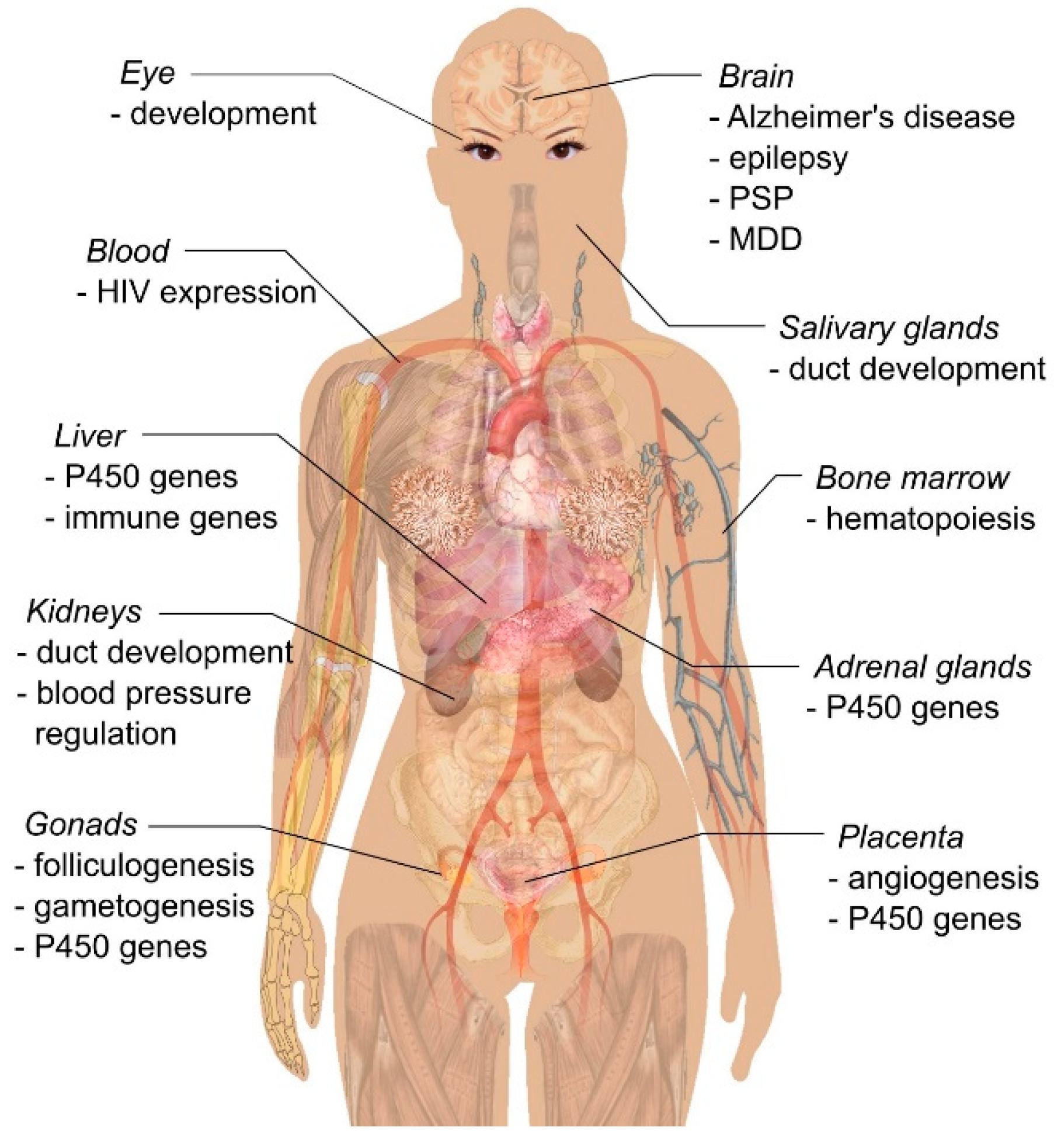
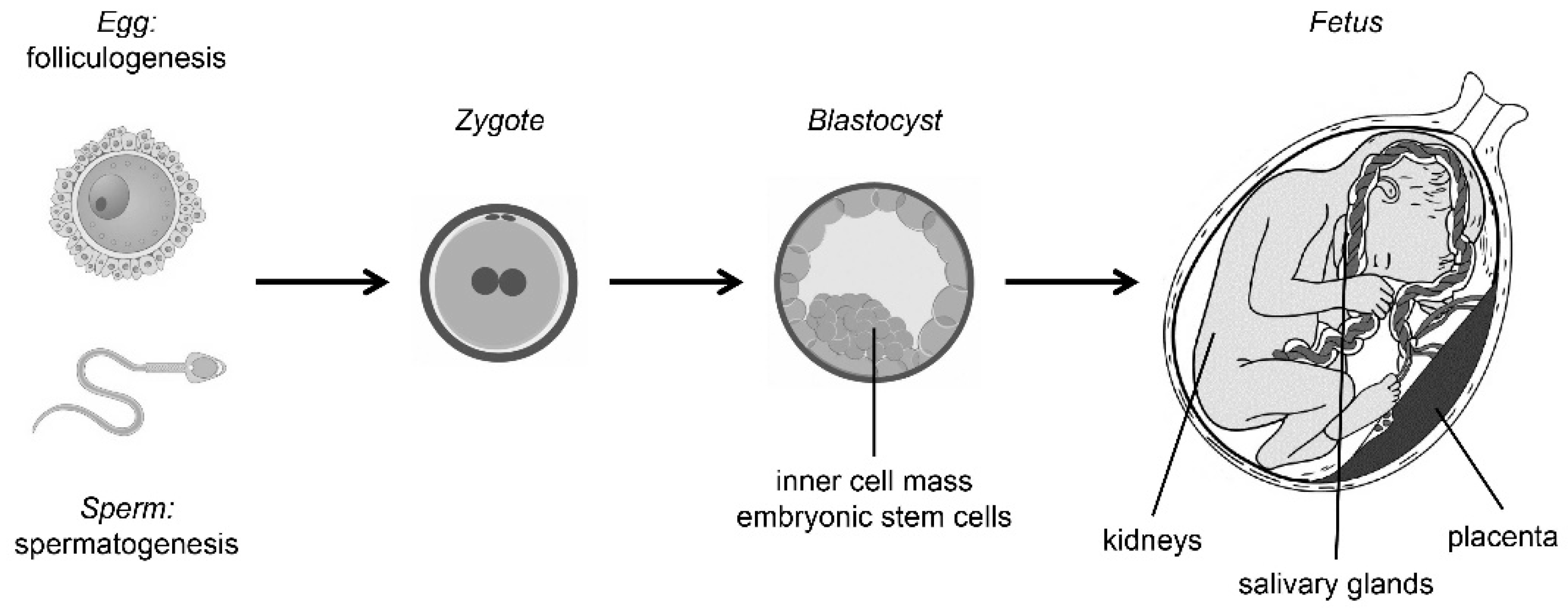
2. Reproduction
3. Embryonic Development
4. Renal Function and Blood-Pressure Regulation
5. Brain Function
6. Other Functions
7. Conclusions and Future Perspectives
Funding
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| AE1 | anion exchanger 1 |
| AQP2 | aquaporin-2 |
| ASIC2a | acid-sensing ion channel 2a |
| ChIP | chromatin immunoprecipitation |
| COX | cyclooxygenase |
| CREB1 | cAMP responsive element binding protein 1 |
| CYP | cytochrome P450 |
| DGCR8 | DiGeorge syndrome critical region gene 8 |
| DSD | disorder of sexual development |
| EMSA | electrophoretic mobility shift assay |
| FGF | fibroblast growth factor |
| GRH | Grainyhead |
| GRHL | Grainyhead-like |
| H+-ATPase | proton-pumping ATPase |
| H+v-ATPase | vacuolar-type H+-transporting ATPase |
| hPGC | human primordial germ cell |
| HS | heparan sulfate |
| HSP | heat shock protein |
| hS/PC | human stem/progenitor cell |
| IC | intercalated cell |
| ICM | inner cell mass |
| INC | intercalating nonciliated cell |
| JAK | Janus kinase |
| KLF | kruppel-like factor |
| LBP | leader-binding protein |
| LH-β | luteinizing hormone β subunit |
| LIF | leukemia inhibitory factor |
| LSF | late SV40 (simian virus 40) factor |
| MAZ | MYC-associated zinc finger protein |
| MDD | major depressive disorder |
| mESC | mouse embryonic stem cell |
| mGC | murine granulosa cell |
| MYF | myogenic factor |
| PAG | pregnancy-associated glycoprotein |
| PC | primordial cell |
| PGE | prostaglandin E |
| PREX | positive regulatory element for xenobiotic response element |
| PSC | proton-secreting cell |
| PSP | progressive supranuclear palsy |
| SF1 | steroidogenic factor 1 |
| SLP | sow lifetime productivity |
| SMG | submandibular gland |
| SNP | single nucleotide polymorphism |
| SOX | SRY-box |
| SRY | sex-determining region Y |
| STAT | signal transducer and activator of transcription |
| TFCP2 | transcription factor cellular promoter-2 |
| TFCP2L1 | TFCP2-like 1 |
| UBP1 | upstream binding protein 1 |
| UTR | untranslated region |
| WNV | West Nile virus |
| YY1 | yin yang 1 |
References
- Traylor-Knowles, N.; Hansen, U.; Dubuc, T.Q.; Martindale, M.Q.; Kaufman, L.; Finnerty, J.R. The evolutionary diversification of LSF and Grainyhead transcription factors preceded the radiation of basal animal lineages. BMC Evol. Biol. 2010, 10, 101. [Google Scholar] [CrossRef] [PubMed]
- Hansen, U.; Owens, L.; Saxena, U.H. Transcription factors LSF and E2Fs: Tandem cyclists driving G0 to S? Cell Cycle 2009, 8, 2146–2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veljkovic, J.; Hansen, U. Lineage-specific and ubiquitous biological roles of the mammalian transcription factor LSF. Gene 2004, 343, 23–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotarba, G.; Krzywinska, E.; Grabowska, A.I.; Taracha, A.; Wilanowski, T. TFCP2/TFCP2L1/UBP1 transcription factors in cancer. Cancer Lett. 2018, 420, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Guo, I.C.; Shih, M.C.; Lan, H.C.; Hsu, N.C.; Hu, M.C.; Chung, B.C. Transcriptional regulation of human CYP11A1 in gonads and adrenals. J. Biomed. Sci. 2007, 14, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.W.; Dietmann, S.; Irie, N.; Leitch, H.G.; Floros, V.I.; Bradshaw, C.R.; Hackett, J.A.; Chinnery, P.F.; Surani, M.A. A Unique Gene Regulatory Network Resets the Human Germline Epigenome for Development. Cell 2015, 161, 1453–1467. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Lei, B.; Li, H.; Zhu, L.; Wang, L.; Tao, H.; Mei, S.; Li, F. MicroRNA-144 is regulated by CP2 and decreases COX-2 expression and PGE2 production in mouse ovarian granulosa cells. Cell Death Dis. 2017, 8, e2597. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Chae, J.H.; Cheon, Y.P.; Kim, C.G. Reciprocal localization of transcription factors YY1 and CP2c in spermatogonial stem cells and their putative roles during spermatogenesis. Acta Histochem. 2016, 118, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.L.; Wang, T.S.; Zhao, M.; Peng, Y.; Fu, Y.S. A Transcriptomic Study of Maternal Thyroid Adaptation to Pregnancy in Rats. Int. J. Mol. Sci. 2015, 16, 27339–27349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bieniek-Kobuszewska, M.; Panasiewicz, G.; Lipka, A.; Majewska, M.; Szafranska, B. Novel SNPs and InDels discovered in two promoter regions of porcine pregnancy-associated glycoprotein 2-like subfamily (pPAG2-Ls) in crossbreed pigs. Funct. Integr. Genom. 2016, 16, 705–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aherrahrou, R.; Aherrahrou, Z.; Kaiser, F.J.; Braunholz, D.; Erdmann, J.; Moumni, M. Identification of a single SNP that affects the promoter activity in the Moroccan prolific D’man breed. J. Anim. Sci. 2015, 93, 2064–2073. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Lee, E.A.; Hong, K.C.; Kim, J.M. Regulatory gene network from a genome-wide association study for sow lifetime productivity traits. Anim. Genet. 2018, 49, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Shinka, T.; Sakamoto, K.; Ewis, A.A.; Nakahori, Y. The male-determining gene SRY is a hybrid of DGCR8 and SOX3, and is regulated by the transcription factor CP2. Mol. Cell. Biochem. 2010, 337, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Ramamurthy, L.; Barbour, V.; Tuckfield, A.; Clouston, D.R.; Topham, D.; Cunningham, J.M.; Jane, S.M. Targeted disruption of the CP2 gene, a member of the NTF family of transcription factors. J. Biol. Chem. 2001, 276, 7836–7842. [Google Scholar] [CrossRef] [PubMed]
- Salamon, S.; Nowacka-Woszuk, J.; Szczerbal, I.; Dzimira, S.; Nizanski, W.; Ochota, M.; Switonski, M. A lack of association between polymorphisms of three positional candidate genes (CLASP2, UBP1, and FBXL2) and canine disorder of sexual development (78,XX; SRY-negative). Sex. Dev. 2014, 8, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Werth, M.; Schmidt-Ott, K.M.; Leete, T.; Qiu, A.; Hinze, C.; Viltard, M.; Paragas, N.; Shawber, C.J.; Yu, W.; Lee, P.; et al. Transcription factor TFCP2L1 patterns cells in the mouse kidney collecting ducts. eLife 2017, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruyama, E.O.; Aure, M.H.; Xie, X.; Myal, Y.; Gan, L.; Ovitt, C.E. Cell-Specific Cre Strains for Genetic Manipulation in Salivary Glands. PLoS ONE 2016, 11, e0146711. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Yonemura, S.; Takada, S. Grainyhead-related transcription factor is required for duct maturation in the salivary gland and the kidney of the mouse. Development 2006, 133, 4737–4748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, P.P.; Patel, V.N.; Liu, S.; Harrington, D.A.; Hoffman, M.P.; Jia, X.; Witt, R.L.; Farach-Carson, M.C.; Pradhan-Bhatt, S. Primary Salivary Human Stem/Progenitor Cells Undergo Microenvironment-Driven Acinar-Like Differentiation in Hyaluronate Hydrogel Culture. Stem Cells Transl. Med. 2017, 6, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Sharpe, P.T.; Miletich, I. Coordination of epithelial branching and salivary gland lumen formation by Wnt and FGF signals. Dev. Biol. 2011, 358, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.N.; Likar, K.M.; Zisman-Rozen, S.; Cowherd, S.N.; Lassiter, K.S.; Sher, I.; Yates, E.A.; Turnbull, J.E.; Ron, D.; Hoffman, M.P. Specific heparan sulfate structures modulate FGF10-mediated submandibular gland epithelial morphogenesis and differentiation. J. Biol. Chem. 2008, 283, 9308–9317. [Google Scholar] [CrossRef] [PubMed]
- Hemphala, J.; Uv, A.; Cantera, R.; Bray, S.; Samakovlis, C. Grainy head controls apical membrane growth and tube elongation in response to Branchless/FGF signalling. Development 2003, 130, 249–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, S.; Li, P.; Tong, C.; Ying, Q.L. Embryonic stem cell self-renewal pathways converge on the transcription factor Tfcp2l1. EMBO J. 2013, 32, 2548–2560. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Forrester-Gauntlett, B.; Turner, P.; Henderson, H.; Oback, B. Signal Inhibition Reveals JAK/STAT3 Pathway as Critical for Bovine Inner Cell Mass Development. Biol. Reprod. 2015, 93, 132. [Google Scholar] [CrossRef] [PubMed]
- Henderson, Y.C.; Frederick, M.J.; Wang, M.T.; Hollier, L.M.; Clayman, G.L. LBP-1b, LBP-9, and LBP-32/MGR detected in syncytiotrophoblasts from first-trimester human placental tissue and their transcriptional regulation. DNA Cell Biol. 2008, 27, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Miller, W.L. Cloning of factors related to HIV-inducible LBP proteins that regulate steroidogenic factor-1-independent human placental transcription of the cholesterol side-chain cleavage enzyme, P450scc. J. Biol. Chem. 2000, 275, 2852–2858. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Miller, W.L. LBP proteins modulate SF1-independent expression of P450scc in human placental JEG-3 cells. Mol. Endocrinol. 2005, 19, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Parekh, V.; McEwen, A.; Barbour, V.; Takahashi, Y.; Rehg, J.E.; Jane, S.M.; Cunningham, J.M. Defective extraembryonic angiogenesis in mice lacking LBP-1a, a member of the grainyhead family of transcription factors. Mol. Cell. Biol. 2004, 24, 7113–7129. [Google Scholar] [CrossRef] [PubMed]
- Ting, S.B.; Wilanowski, T.; Cerruti, L.; Zhao, L.L.; Cunningham, J.M.; Jane, S.M. The identification and characterization of human Sister-of-Mammalian Grainyhead (SOM) expands the grainyhead-like family of developmental transcription factors. Biochem. J. 2003, 370, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Walentin, K.; Hinze, C.; Werth, M.; Haase, N.; Varma, S.; Morell, R.; Aue, A.; Potschke, E.; Warburton, D.; Qiu, A.; et al. A Grhl2-dependent gene network controls trophoblast branching morphogenesis. Development 2015, 142, 1125–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilanowski, T.; Tuckfield, A.; Cerruti, L.; O’Connell, S.; Saint, R.; Parekh, V.; Tao, J.; Cunningham, J.M.; Jane, S.M. A highly conserved novel family of mammalian developmental transcription factors related to Drosophila grainyhead. Mech. Dev. 2002, 114, 37–50. [Google Scholar] [CrossRef]
- Tchapyjnikov, D.; Li, Y.; Pisitkun, T.; Hoffert, J.D.; Yu, M.J.; Knepper, M.A. Proteomic profiling of nuclei from native renal inner medullary collecting duct cells using LC-MS/MS. Physiol. Genom. 2010, 40, 167–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koutnikova, H.; Laakso, M.; Lu, L.; Combe, R.; Paananen, J.; Kuulasmaa, T.; Kuusisto, J.; Haring, H.U.; Hansen, T.; Pedersen, O.; et al. Identification of the UBP1 locus as a critical blood pressure determinant using a combination of mouse and human genetics. PLoS Genet. 2009, 5, e1000591. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L. Molecular biology of steroid hormone synthesis. Endocr. Rev. 1988, 9, 295–318. [Google Scholar] [CrossRef] [PubMed]
- Morohashi, K.; Sogawa, K.; Omura, T.; Fujii-Kuriyama, Y. Gene structure of human cytochrome P-450(SCC), cholesterol desmolase. J. Biochem. 1987, 101, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Nebert, D.W.; Nelson, D.R.; Coon, M.J.; Estabrook, R.W.; Feyereisen, R.; Fujii-Kuriyama, Y.; Gonzalez, F.J.; Guengerich, F.P.; Gunsalus, I.C.; Johnson, E.F.; et al. The P450 superfamily: Update on new sequences, gene mapping, and recommended nomenclature. DNA Cell Biol. 1991, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, M.; Walkowska, A.; Mlacki, M.; Pistolic, J.; Wrzesinski, T.; Benes, V.; Jane, S.M.; Wesoly, J.; Kompanowska-Jezierska, E.; Wilanowski, T. Consequences of the loss of the Grainyhead-like 1 gene for renal gene expression, regulation of blood pressure and heart rate in a mouse model. Acta Biochim. Pol. 2015, 62, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Walkowska, A.; Pawlak, M.; Jane, S.M.; Kompanowska-Jezierska, E.; Wilanowski, T. Effects of high and low sodium diet on blood pressure and heart rate in mice lacking the functional grainyhead-like 1 gene. Physiol. Res. 2017, 66, 163–165. [Google Scholar] [PubMed]
- Quigley, I.K.; Stubbs, J.L.; Kintner, C. Specification of ion transport cells in the Xenopus larval skin. Development 2011, 138, 705–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsura, A.; Kimura, K.; Hosoi, K.; Tomokuni, Y.; Nesori, M.; Goryo, K.; Numayama-Tsuruta, K.; Torii, S.; Yasumoto, K.; Gotoh, O.; et al. Transactivation activity of LBP-1 proteins and their dimerization in living cells. Genes Cells 2009, 14, 1183–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, F.; Yasumoto, K.; Kimura, K.; Numayama-Tsuruta, K.; Sogawa, K. Heterodimerization with LBP-1b is necessary for nuclear localization of LBP-1a and LBP-1c. Genes Cells 2005, 10, 861–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- To, S.; Rodda, S.J.; Rathjen, P.D.; Keough, R.A. Modulation of CP2 family transcriptional activity by CRTR-1 and sumoylation. PLoS ONE 2010, 5, e11702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, J.B.; Li, G.; Roeder, R.G. Characterization of a family of related cellular transcription factors which can modulate human immunodeficiency virus type 1 transcription in vitro. Mol. Cell. Biol. 1994, 14, 1776–1785. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Gao, G.; Zhang, Y.; Sun, Y.; Li, H.; Dong, S.; Ma, W.; Liu, B.; Wang, W.; Wu, H.; et al. Glucose Deficiency Elevates Acid-Sensing Ion Channel 2a Expression and Increases Seizure Susceptibility in Temporal Lobe Epilepsy. Sci. Rep. 2017, 7, 5870. [Google Scholar] [CrossRef] [PubMed]
- Rademakers, R.; Melquist, S.; Cruts, M.; Theuns, J.; Del-Favero, J.; Poorkaj, P.; Baker, M.; Sleegers, K.; Crook, R.; De Pooter, T.; et al. High-density SNP haplotyping suggests altered regulation of tau gene expression in progressive supranuclear palsy. Hum. Mol. Genet. 2005, 14, 3281–3292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zubenko, G.S.; Hughes, H.B., 3rd. Effects of the G(-656)A variant on CREB1 promoter activity in a glial cell line: Interactions with gonadal steroids and stress. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2008, 147, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Zubenko, G.S.; Hughes, H.B., 3rd; Stiffler, J.S.; Brechbiel, A.; Zubenko, W.N.; Maher, B.S.; Marazita, M.L. Sequence variations in CREB1 cosegregate with depressive disorders in women. Mol. Psychiatry 2003, 8, 611–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, K.F.; Miller, C.C.; Anderton, B.H.; Shaw, P.C. Molecular cloning and characterization of the human glycogen synthase kinase-3beta promoter. Genomics 1999, 60, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.V.; Budd Haeberlein, S.L.; Avila, J. Glycogen synthase kinase 3: A drug target for CNS therapies. J. Neurochem. 2004, 89, 1313–1317. [Google Scholar] [CrossRef] [PubMed]
- Jope, R.S.; Yuskaitis, C.J.; Beurel, E. Glycogen synthase kinase-3 (GSK3): Inflammation, diseases, and therapeutics. Neurochem. Res. 2007, 32, 577–595. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.C.; Goumidi, L.; Vrieze, F.W.; Frigard, B.; Harris, J.M.; Cummings, A.; Coates, J.; Pasquier, F.; Cottel, D.; Gaillac, M.; et al. The transcriptional factor LBP-1c/CP2/LSF gene on chromosome 12 is a genetic determinant of Alzheimer’s disease. Hum. Mol. Genet. 2000, 9, 2275–2280. [Google Scholar] [CrossRef] [PubMed]
- Schahab, S.; Heun, R.; Schmitz, S.; Maier, W.; Kolsch, H. Association of polymorphism in the transcription factor LBP-1c/CP2/LSF gene with Alzheimer’s disease and major depression. Dement. Geriatr. Cogn. Disord. 2006, 22, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Imabuchi, R.; Ohmiya, Y.; Kwon, H.J.; Onodera, S.; Kitamura, N.; Kurokawa, T.; Gong, J.P.; Yasuda, K. Gene expression profile of the cartilage tissue spontaneously regenerated in vivo by using a novel double-network gel: Comparisons with the normal articular cartilage. BMC Musculoskelet. Disord. 2011, 12, 213. [Google Scholar] [CrossRef] [PubMed]
- Caddy, J.; Wilanowski, T.; Darido, C.; Dworkin, S.; Ting, S.B.; Zhao, Q.; Rank, G.; Auden, A.; Srivastava, S.; Papenfuss, T.A.; et al. Epidermal wound repair is regulated by the planar cell polarity signaling pathway. Dev. Cell 2010, 19, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Ting, S.B.; Caddy, J.; Hislop, N.; Wilanowski, T.; Auden, A.; Zhao, L.L.; Ellis, S.; Kaur, P.; Uchida, Y.; Holleran, W.M.; et al. A homolog of Drosophila grainy head is essential for epidermal integrity in mice. Science 2005, 308, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Mace, K.A.; Pearson, J.C.; McGinnis, W. An epidermal barrier wound repair pathway in Drosophila is mediated by grainy head. Science 2005, 308, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, M.; Mukesh, M.; Kishore, A.; Mishra, B.P.; Kataria, R.S.; Joshi, B.K. Novel polymorphisms in UTR and coding region of inducible heat shock protein 70.1 gene in tropically adapted Indian zebu cattle (Bos indicus) and riverine buffalo (Bubalus bubalis). Gene 2013, 527, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Trung, N.T.; Engelke, R.; Mittler, G. SILAC-Based Quantitative Proteomics Approach to Identify Transcription Factors Interacting with a Novel Cis-Regulatory Element. J. Proteom. Bioinform. 2014, 7, 82–87. [Google Scholar]
- Kurose, K.; Tohkin, M.; Hasegawa, R. Transcription factor NF2d9 (LBP-1a) interacts with the positive regulatory element for the xenobiotic responsive element. Biochim. Biophys. Acta 2005, 1727, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Sueyoshi, T.; Kobayashi, R.; Nishio, K.; Aida, K.; Moore, R.; Wada, T.; Handa, H.; Negishi, M. A nuclear factor (NF2d9) that binds to the male-specific P450 (Cyp 2d-9) gene in mouse liver. Mol. Cell. Biol. 1995, 15, 4158–4166. [Google Scholar] [CrossRef] [PubMed]
- Hecker, M.; Goertsches, R.H.; Fatum, C.; Koczan, D.; Thiesen, H.J.; Guthke, R.; Zettl, U.K. Network analysis of transcriptional regulation in response to intramuscular interferon-beta-1a multiple sclerosis treatment. Pharmacogenom. J. 2012, 12, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Long, D.; Deng, X.; Singh, P.; Loeb, M.; Lauring, A.S.; Seielstad, M. Identification of genetic variants associated with susceptibility to West Nile virus neuroinvasive disease. Genes Immun. 2016, 17, 298–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, S.; Yang, Z.; Du, L.; Jiang, Z.; Shu, Q.; Chen, Y.; Li, F.; Zhou, Q.; Ohno, S.; Chen, R.; et al. Identification of a susceptibility locus in STAT4 for Behcet’s disease in Han Chinese in a genome-wide association study. Arthritis Rheum. 2012, 64, 4104–4113. [Google Scholar] [CrossRef] [PubMed]
- Kokoszynska, K.; Ostrowski, J.; Rychlewski, L.; Wyrwicz, L.S. The fold recognition of CP2 transcription factors gives new insights into the function and evolution of tumor suppressor protein p53. Cell Cycle 2008, 7, 2907–2915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donaghey, J.; Thakurela, S.; Charlton, J.; Chen, J.S.; Smith, Z.D.; Gu, H.; Pop, R.; Clement, K.; Stamenova, E.K.; Karnik, R.; et al. Genetic determinants and epigenetic effects of pioneer-factor occupancy. Nat. Genet. 2018, 50, 250–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavi, O. Redundancy: A critical obstacle to improving cancer therapy. Cancer Res. 2015, 75, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Cereda, M.; Mourikis, T.P.; Ciccarelli, F.D. Genetic Redundancy, Functional Compensation, and Cancer Vulnerability. Trends Cancer 2016, 2, 160–162. [Google Scholar] [CrossRef] [PubMed]
- Koehler, A.N. A complex task? Direct modulation of transcription factors with small molecules. Curr. Opin. Chem. Biol. 2010, 14, 331–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurlak, L.O.; Knofler, M.; Mistry, H.D. Lumps & Bumps: Common features between placental development and cancer growth. Placenta 2017, 56, 2–4. [Google Scholar] [PubMed]
- Wang, Q.; Lash, G.E. Angiopoietin 2 in placentation and tumor biology: The yin and yang of vascular biology. Placenta 2017, 56, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Haider, S.; Pollheimer, J.; Knofler, M. Notch signalling in placental development and gestational diseases. Placenta 2017, 56, 65–72. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taracha, A.; Kotarba, G.; Wilanowski, T. Neglected Functions of TFCP2/TFCP2L1/UBP1 Transcription Factors May Offer Valuable Insights into Their Mechanisms of Action. Int. J. Mol. Sci. 2018, 19, 2852. https://doi.org/10.3390/ijms19102852
Taracha A, Kotarba G, Wilanowski T. Neglected Functions of TFCP2/TFCP2L1/UBP1 Transcription Factors May Offer Valuable Insights into Their Mechanisms of Action. International Journal of Molecular Sciences. 2018; 19(10):2852. https://doi.org/10.3390/ijms19102852
Chicago/Turabian StyleTaracha, Agnieszka, Grzegorz Kotarba, and Tomasz Wilanowski. 2018. "Neglected Functions of TFCP2/TFCP2L1/UBP1 Transcription Factors May Offer Valuable Insights into Their Mechanisms of Action" International Journal of Molecular Sciences 19, no. 10: 2852. https://doi.org/10.3390/ijms19102852