The Role of Tumor-Associated Neutrophils in Colorectal Cancer


Department of Surgery, Graduate School of Medicine, Kyoto University, Kyoto 606-8507, Japan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(3), 529; https://doi.org/10.3390/ijms20030529
Submission received: 28 December 2018
/
Revised: 24 January 2019
/
Accepted: 24 January 2019
/
Published: 27 January 2019
(This article belongs to the Special Issue Basic and Translational Research in Colorectal Cancer)
Abstract
:Colorectal cancer (CRC) is one of the most common causes of cancer deaths worldwide and the number of CRC patients is increasing progressively. Despite the improvement of the surgical techniques and chemotherapy, we have not completely overcome this disease yet due to the metastases. Therefore, understanding the mechanisms through which metastasis occurs is important for overcoming CRC. Normal host cells in the tumor microenvironment, such as macrophages and fibroblasts, have been reported to promote the growth of CRCs. Although neutrophils were originally considered to have defensive functions against tumor cells, it has been revealed that some populations of neutrophils, called as tumor-associated neutrophils (TANs), have tumor-supportive functions. The plasticity between tumor-suppressive and -supportive neutrophils are regulated by transforming growth factor (TGF)-β and Interferon-β signaling. Some studies have demonstrated that TANs promote the spread of cancer cells to distant organs. TANs contribute to the tumor invasion and angiogenesis through the production of matrix metalloproteinase-9 (MMP9), vascular endothelial growth factor (VEGF), and hepatocyte growth factor (HGF) in the primary and metastatic sites. Neutrophils also promotes tumor cell dissemination by capturing circulating tumor cells using neutrophil extracellular traps and promote their migration to distant sites. The neutrophil-to-lymphocyte ratio is a well-defined predictive marker for CRC patients. In this review, we highlight the molecular signaling between TANs and CRC cells and the possibility of TANs as a potential target for cancer therapy.
1. Introduction
Colorectal cancer (CRC) is one of the most common causes of cancer-related deaths worldwide [1,2,3]. Despite advances in surgical techniques, chemo-drugs, and molecular-targeted drugs (e.g., bevacizumab and cetuximab targeting vascular endothelial growth factor (VEGF) and epidermal growth factor receptor (EGFR), respectively) [4], the number of CRC patients is increasing progressively [5,6]. At least one third of CRC patients develop liver metastases, and CRC-related death is usually attributable to distant metastasis [7,8]. Once the disease spreads to distant organs, neither conventional chemotherapy nor current targeted therapy offers significant benefits. Therefore, it is important to understand the mechanisms through which metastasis occurs and to find therapeutic targets for distant metastasis.
The process of metastatic formation can be divided into several successive steps (Figure 1). In the primary tumor site, the transformed tumor cells begin to grow and secrete angiogenic factors, which results in extensive vascularization. Tumor cells locally invade through the activation of proteases and intravasate into thin-walled vessels (i.e., venules and lymphatic vessels) and enter the blood circulation. Embolization of single cancer cell or aggregates occur next. During this process, most circulating cancer cells are destroyed by the shear forces of blood flow or by the attack from components of the host immune system such as natural killer cells. If the tumor cells can survive in blood circulation, they become trapped in the capillary beds of distant organs. Finally, tumor cells extravasate into the organ parenchyma and start to form micrometastases. Some tumor cells within micrometastatic sites die due to the attack of host immune cells, while others survive in a dormant state that exits from the cell cycle and balances their proliferation and apoptosis. Although less is understood about how dormancy is broken, some tumor cells start to proliferate and expand through the secretion of angiogenic factors and the activation of proteases to form metastatic colonies. Only a limited number of cancer cells can form metastases in distant organs [9,10]. The transition from pre-angiogenic to angiogenic metastasis is a rate-limiting step in the occurrence of liver metastasis, which suggests that the development of an angiogenic phenotype is a key step for metastatic progression [11].
However, the precise underlying mechanisms by which cancer cells survive in the hostile environment and develop metastatic sites still remain unclear. It has been reported that several types of host cells, such as fibroblasts (cancer-associated fibroblasts: CAF), macrophages (tumor-associated macrophages: TAMs), and mesenchymal stem cells, play important roles in the formation of the tumor microenvironment [12,13,14]. In addition, recent accumulating evidence has shown that some populations of neutrophils, known as tumor-associated neutrophils (TANs), could support the growth, invasion, and angiogenesis of cancer cells, although they have been classically considered to exhibit a defensive response against tumor cells. They have also been reported to exert supportive functions in the development of metastasis. Here, we highlight the role of TANs in supporting the development of distant CRC metastasis, especially liver metastasis.
Liver metastasis is a complex, multistep process. In the primary tumor site, transformed tumor cells start to proliferate and secrete angiogenic factors, which results in extensive vascularization. Tumor cells locally invade blood vessels. Most circulating tumor cells are destroyed by the shear forces of blood flow or by the attack from the host immune system such as natural killer cells. If the tumor cells can survive in blood circulation, they become trapped in the capillary beds of distant organs. Finally, tumor cells extravasate into the organ parenchyma and start to form micrometastases. Some tumor cells die and others survive in a dormant state. Some tumor cells break the dormancy and start to proliferate and expand through the secretion of angiogenic factors and the activation of proteases.
2. Tumor-Associated Neutrophils (TANs)
Normal host cells in the tumor microenvironment, such as CAFs and TAMs, assist in the growth, invasion, and metastasis of cancer cells [13,14]. It has become evident that bone marrow-derived cells including TAMs, TANs, and myeloid-derived suppressor cells (MDSCs), contribute to tumor progression [12,15,16,17,18,19]. Recently, a number of studies have demonstrated through immunohistochemical analyses that neutrophils, which are another leukocyte population, were intermingled in various cancer tissues. Chemokines are small peptides binding to G protein-coupled receptors to induce chemoattraction, inflammation, and/or angiogenesis [20]. They are one of the key factors that facilitate cancer metastasis [21]. Tumor cells often produce several inflammatory chemokines, including neutrophil-attracting CXC-chemokines [22,23]. The migration of neutrophils toward the tumor is mainly mediated by CXC-chemokines that bind to CXCR1 and/or CXCR2 [24,25,26].
Neutrophils have been originally viewed as the first-responder of the innate immune system in the resistance against extracellular pathogens. However, recent evidence has added a new aspect on the function of neutrophils. Neutrophils are involved in the regulation of innate and adaptive immune systems, and can be polarized towards distinct phenotypes in response to environmental signals [22]. They are classically characterized based on their ability to induce phagocytosis, release lytic enzymes, and produce reactive oxygen species (ROS) [27,28]. In the context of the tumor microenvironment, accumulating evidence has revealed the prominent role of neutrophils in infiltrating tumor tissues to promote their growth, invasion, angiogenesis, and metastasis in various types of cancers, although they were initially considered to have a defensive function against tumor cells [29,30,31].
TAMs are divided into two populations: the anti-tumorigenic “M1” phenotype and pro-tumorigenic “M2” phenotype. As with TAMs, recent studies have suggested that TANs also exhibit considerable plasticity and are capable of polarization into either an anti-tumorigenic “N1” phenotype or a pro-tumorigenic “N2” phenotype [22,32,33]. Their surface markers, transcriptional regulators, and cytokine profiles remain to be investigated. Neutrophils are known to secrete several inflammatory, immunoregulatory, and angiogenic factors, including neutrophil elastase [34], matrix metalloproteinases (MMPs), vascular endothelial growth factor (VEGF) [35,36], and hepatocyte growth factor [37], which can exhibit paracrine effects on the tumor microenvironment. “N1” neutrophils exhibit increased cytotoxicity and reduced immunosuppressive ability by the production of tumor necrosis factor (TNF)-α, intercellular adhesion molecule (ICAM)-1, ROS, and Fas and by decreasing arginase expression. In contrast, “N2” neutrophils support tumor expansion by expressing arginase, MMP-9, VEGF, and numerous chemokines including CCL2, CCL5 and CXCL4 [32]. Fridlender et al. reported that transforming growth factor (TGF)-β signaling functions as a regulator between the N1 and N2 phenotypes. TGF-β within tumors skews differentiation toward the N2 phenotype, while inhibition of TGF-β signaling induces an anti-tumoral N1 phenotype [32]. Interferon-β was also recently reported to induce an N1 phenotype [38]. Taken together, the phenotype of TANs depends on the signals encountered in the tumor microenvironment.
Moreover, some studies have recently investigated the possible involvement of neutrophil extracellular traps (NETs) in promoting the migration and extravasation of cancer cells. NETs are composed mainly of fibers of decondensed DNA, and are decorated with proteins released from activated neutrophils [39,40,41,42,43]. They act as meshes that trap microorganisms and, in turn, promote the interaction between pathogens and neutrophil-derived effector molecules [39,44]. NETs have recently been suggested to capture circulating cancer cells and promote their migration to new sites [45,46]. NETs has also been shown that they can activate toll-like receptor 9 on CRC cells, resulting in cellular growth, migration, and invasion via activation of mitogen-activated protein kinase (MAPK) signaling [47]. Najmeh et al. recently reported the importance of β1-integrin expression on both circulating cancer cells and NETs in mediating cancer cell adhesion to NETs in vivo, resulting in the development of metastatic diseases [48].
In addition, TAMs and TANs have the potential to drive tumor angiogenesis. In various murine models, TAMs and TANs are major sources of MMP9 [49], which promotes angiogenesis by its extracellular matrix-degrading properties [50]. In a genetically-engineered mouse model of pancreatic cancer, MMP9 expression was exclusively found in neutrophils, and neutrophil depletion inhibited the angiogenic switch in the primary tumors [51]. Moreover, in a tumor xenograft model, granulocyte colony-stimulating factor (G-CSF)-induced upregulation of Bv8 (known as prokineticin-2) in neutrophil was shown to promote tumor angiogenesis [52]. G-CSF facilitated neutrophil recruitment into the tumor, stimulated Bv8 expression, and promoted angiogenesis, which resulted in resistance against anti-VEGF treatment [53,54,55].
CXCR2 and its ligands (i.e., CXCL1, CXCL2, CXCL3, CXCL5, CXCL7, and CXCL8) are responsible for the recruitment of neutrophils under normal physiological conditions and are implicated in the mobilization of TANs [56]. In tumor-bearing mouse models, targeting CXCR2-mediated TAN mobilization has been reported to increase the number of tumor-infiltrating lymphocytes and potentiate anti-programmed death 1 checkpoint blockade [57,58,59,60]. Nywening et al. have recently reported that dual blocking of CXCR2+ TANs and CCR2+ TAMs disrupted the recruitment of myeloid cells and improved anti-tumor immunity in a mouse model of pancreatic cancer [61]. Recently, we have found that loss of SMAD4 from CRC cells results in the secretion of CXCL1 and CXCL8 to recruit CXCR2+ neutrophils, and that, in turn, the recruited neutrophils abundantly produce CXCL1 and CXCL8, which further prompts the accumulation of CXCR2+ neutrophils and results in an amplification of the cytokine/chemokine milieu shaped by the CXCL1/8-CXCR2 axis [62].
3. TANs in Human CRC
The relationship between TAN infiltration and human cancer prognosis has not been systemically investigated, although some studies have reported the role of neutrophils as a prognostic factor in various types of human cancers. The increase of neutrophil count in peripheral blood (i.e., neutrophil-to-lymphocyte ratio (NLR)) has been shown to be related to poor clinical outcomes in pancreatic cancer, gastric cancer, and breast cancer [63,64,65], emphasizing the importance of neutrophils in cancer biology. High NLR has also been demonstrated as a poor prognostic factor in CRC patients. Li et al. retrospectively analyzed a cohort of 354 CRC patients with stage I–III cancer and revealed a strong relationship between dynamic changes in NLR and overall survival [66]. Other studies have also reported that high NLR had an adverse effect on overall survival in CRC patients subjected to curative surgery [67,68]. High NLR was also shown to predict poor outcome following hepatic resection for liver metastasis of CRC [69]. Dell’Aquila et al. showed that high NLR was also a poor prognostic factor in unresectable metastatic CRC patients treated with bevacizumab plus chemotherapy [70].
However, the effect of intratumoral neutrophils on the survival for CRC patients is still unclear. Rao et al. demonstrated that the increase in intratumoral neutrophils was associated with malignant phenotypes and could predict adverse prognosis in CRC [71]. On the other hand, Berry et al. analyzed the number of neutrophils in CRC tissues by counting neutrophils manually based on their morphology because of the lack of neutrophil-specific antibodies and demonstrated that high levels of TANs were associated with improved overall survival in patients with stage II CRC [72]. As described, the signals from the tumor microenvironment that determine the N1 and N2 neutrophil phenotypes might affect the results. We have recently reported that loss of SMAD4-promoted CCL15 expression from CRC cells to recruit CCR1+ myeloid cells through the CCL15-CCR1 axis, and that CCL15 expression in primary and metastatic CRCs was a predictor of CRC patients’ prognosis [73,74,75]. Most CCR1+ myeloid cells recruited into the primary CRC and metastatic CRC were of the granulocytic-MDSC phenotype (CD11b+, CD33+, HLA-DR−, and CD15+) [74] and TAN phenotype (CD11b+, CD33−, HLA-DR−, CD15+, and CD16+) [75], respectively. MDSCs constitute a heterogeneous population of immature myeloid cells at various differentiation stages and represent a group of myeloid cells that suppress immune responses. Because TANs and MDSCs share a common set of markers and are morphologically similar, it remains unclear whether TANs and MDSCs are separate populations or not [29,76,77]. In addition, MDSCs were reported to differentiate into mature TAMs or TANs within the tumor [64]. Further analysis is needed to evaluate the significance of intratumoral neutrophils in CRC.
4. TANs in Animal Model for CRC Liver Metastasis
A number of studies have tried to clarify the underlying mechanisms by which neutrophils support liver metastasis of CRC cells in vitro and in vivo using metastatic mouse models (Table 1).
In a mouse model of liver metastasis in which tumor cells were inoculated through splenic injection, Hirai et al. demonstrated the interaction between neutrophils and CRC cells during the process of colonization within the liver [78]. They found that mouse CCL9 (mCCL9)-expressing CRC cells recruited myeloid cells expressing CCR1, the cognate receptor of mCCL9, to form early metastatic foci in the liver, and that four distinct types of myeloid cells were recruited to the site of liver metastasis: CCR1+ neutrophils, monocytes, eosinophils, and fibrocytes. CCR1+ neutrophils produce MMP9, which helps cancer foci to expand in the early stage, and trigger the recruitment of fibrocytes and monocytes that produce MMP2 during the later stages. We have also reported that the recruitment of CCR1+ myeloid cells facilitates primary CRC invasion [74] and metastasis to the liver [73] and lungs [75]. Using Ccr1-knockout mice, Rodero et al. reported that CCR1 expression in hematopoietic and non-hematopoietic cells facilitated liver metastasis through myeloid cell accumulation in the metastatic tumors [79]. Moreover, using the bone-marrow transplantation model of Ccr1-knockout mice, we have recently found that knockout of CCR1 expression in myeloid cells significantly suppressed tumor growth in primary and metastatic CRCs [80], which suggests that the use of CCR1 inhibitors can be a promising strategy to treat CRC.
CXCL8, or interleukin(IL)-8, is the first-described angiogenic chemokine, and is secreted from CRC cells stimulated by TNF-α and IL-1α [88]. CXCL8 is also induced by hypoxia as a hypoxia-inducible factor 1-independent pathway to preserve tumor angiogenesis as a compensatory pathway of VEGF [89]. Kumar et al. revealed that the upregulation of CXCL8 secreted from CRC cells promoted CRC liver metastasis [81]. They demonstrated that the expression of CXCL8 was upregulated in the invasion front of the tumor and that shRNA-mediated knockdown of CXCL8 resulted in significantly decreased cell proliferation, migration, and invasion in vitro and dramatic reduction of tumor metastasis in vivo. They also revealed that the knockdown of CXCL8 resulted in a reduction in VEGF-A expression, suggesting that overexpression of CXCL8 could induce a VEGF-dependent angiogenic response. CXCL8 could also promote angiogenesis in a VEGF-independent manner in CRC [90]. Using a CRC xenograft model, Yamamoto et al. reported that the CXCL1-CXCR2 axis was important in modulating the pre-metastatic niche of CRC liver metastasis, and that TSU68 (an inhibitor of VEGF receptor 2, platelet-derived growth factor receptor β and fibroblast growth factor (FGF) receptor 1) suppressed CXCL1 expression in the pre-metastatic liver, resulting in suppression of CXCR2+ neutrophils homing and subsequent liver metastasis [82]. In Cxcr2-knockout mice, the deficiency of the CXCL8-CXCR2 axis in the host cells resulted in the inhibition of CRC growth and metastasis [91]. Moreover, the CXCR1/CXCR2 antagonist inhibited CRC liver metastasis by decreasing tumor angiogenesis and facilitating tumor cell apoptosis in a mouse model [83]. Combination of a CXCR2 antagonist and oxaliplatin was reported to result in a great decrease of tumor growth and angiogenesis in xenograft models [92]. Using CXCL8-expressing transgenic mice, Asfaha et al. reported that CXCL8 expression increased the mobilization of immature myeloid cells in dextran sodium sulfate-induced colitis, which exacerbated acute inflammation and accelerated colon carcinogenesis [93].
Other signaling molecules are indicated to be involved in the pro-tumorigenic function of neutrophils. Gordon-Weeks et al. demonstrated that human CRC metastasis to the liver and experimental murine models of liver metastasis were infiltrated by neutrophils. They showed that metastasis-associated neutrophils in the liver substantially expressed FGF 2, a pro-angiogenic molecule, indicating neutrophil polarization by the tumor microenvironment [84]. Of note, neutralizing anti-FGF2 antibodies could cause neutrophil depletion and reduce liver metastatic colony growth and vascular density.
Some studies have revealed that signaling pathways in the liver itself make it more susceptible to metastasis and regulate the polarization of the TAN phenotype. In transgenic mice with a conditional, liver-specific insulin-like growth factor (IGF)-1 deficiency, Rayes et al. showed that IGF-1 signaling in the liver was important in the polarization of neutrophils. In mice subjected to IGF-1 deficiency three weeks but not two days prior to the inoculation of CRC cells, infiltrated neutrophils in the liver did not show characteristics of tumor-promoting phenotypes, although the number of neutrophils was increased. They suggested that sustained IGF-1 deficiency is necessary to alter the neutrophil phenotype [85].
Seubert et al. demonstrated that high systemic expression of tissue inhibitor of metalloproteases-1 (TIMP-1) increased liver susceptibility towards metastasis by triggering the formation of a pre-metastatic niche. High systemic levels of TIMP-1 resulted in increased hepatic CXCL12 levels, which in turn promoted recruitment of neutrophils to the liver. Both inhibition of CXCL12-dependent neutrophil recruitment and systemic depletion of neutrophils could suppress TIMP-1-induced susceptibility towards liver metastasis [86].
Using a colitis-associated CRC mouse model, Ma et al. reported that prostaglandin E2 receptor subtype EP2 was expressed in infiltrating TANs and CAFs in CRC, and that the expression of cytokines such as TNF-α, IL-6, CXCL1, cyclooygenase-2, and Wnt5A was amplified in tumor lesions via EP2 expression in TANs and CAFs. Importantly, treatment with a selective EP2 antagonist potently suppressed tumor growth in this model [87].
5. Limitations of Studies on the Interaction Between Neutrophils and Cancer Cells
In most studies, the interaction between neutrophils and CRC cells has been analyzed in vitro using cancer cells, isolated neutrophils and human umbilical vein endothelial cells. However, isolated neutrophils for in vitro experiments do not behave normally as they are primed or pre-activated during the process of isolation [94]. In addition, some studies with in vivo experiments use xenograft models to show the interaction between neutrophils and CRC cells. However, xenograft models are less reliable for cancer metastasis. Therefore, we must keep in mind that the mechanisms that are observed through in vitro experiments or in vivo experiments with xenografts do not always represent actual in vivo biological phenomena. Although the development of microscopic techniques, such as confocal microscopy and two-photon excitation microscopy, combined with fluorescent proteins have enabled us to visualize various biological events in vivo, it is still challenging to observe the interaction between cancer cells and neutrophils during the process of liver metastasis. Further in vivo studies will be required.
Accumulating evidence indicated that exogenous and endogenous factors, such as diet, alcohol, smoking, obesity, lifestyle, environmental exposures, and microbiome, can influence the tumor–immune interactions. Through recruitment of host immune cells, the gut microbiome could generate a proinflammatory microenvironment that is conductive for CRC progression [95]. Molecular pathological epidemiology (MPE) integrates tumor immunology into population health sciences, and links the exposures and germline genetics to tumor and immune characteristics using bioinformatics, in vivo pathology and omics technologies. This kind of integrative approach would be important to understand the mechanisms of tumor progression, effective prevention and therapeutic strategies for precision medicine for CRC [96,97].
6. TANs as a Potential Target for Cancer Therapy
The evaluation of TANs as a potential therapeutic target is still ongoing because their role in cancer development and metastasis is not completely understood. Considering the role of TANs in tumor progression, targeting neutrophils in cancer could be a potential new anti-tumor therapy. However, depletion of neutrophils in humans could lead to self-defeating immunosuppression as neutrophils are essential for host defense against infection. Therefore, it has been postulated that blocking specific populations of neutrophils, especially TANs, can be beneficial and promote tumor regression or metastatic spread.
Since TGF-β modulates the pro- and anti-tumor phenotypes of neutrophils, TGF-β blocking could theoretically be a potential therapeutic strategy. Multiple trials that have tested the effect of TGF-β blocking failed as a result of significant side effects because TGF-β is involved in numerous physiological pathways [98]. New strategies and molecules directed toward either TGF-β or its receptors are currently being clinically tested [99]. In CRC, a TGF-β receptor II antibody (IMC-TR1, also known as LY3022859) has been developed, and the murinized derivative exhibited good response in mouse models of CRC and breast cancer [100]. At the time of writing, this drug is in a phase I trial for patients with advanced solid tumors including CRC for whom standard therapies have failed (NCT01646203) [101].
Chemokine blocking could be another effective strategy resulting in impaired neutrophil recruitment to the tumor. Since CXCL8 secreted from CRC cells recruits neutrophils [81,88,89], blocking the CXCL8 axis by neutralizing antibodies could be a good therapeutic approach. However, its consequences on the phenotype of circulating or intratumoral neutrophils in human cancer are still unknown. Therefore, further studies are needed to gain a detailed understanding of TANs and CRC and for the application of future novel anti-tumor therapies.
7. Conclusions
Accumulating evidence has shown that neutrophils infiltrating CRC tissues, as well as macrophages and fibroblasts, play important roles in the tumor microenvironment. TANs exhibit the plasticity between the anti-tumorigenic N1 or tumor-promoting N2 phenotype, which is determined by signals from surrounding tissues. In this review, we highlighted the role of neutrophils in promoting liver metastasis of CRC. NLR is a well-defined predictive biomarker for CRC patients. Studies with animal models for liver metastasis of CRC demonstrated the underlying mechanisms by which neutrophils promoted liver metastasis, which could contribute to novel therapeutic targets and biological markers.
Author Contributions
R.M. and K.K. wrote the manuscript. R.M. and K.K. contributed the conception and design of the manuscript. Y.I., R.O., Y.K., and Y.S. reviewed the manuscript for important intellectual content.
Funding
This research was funded by grants from the Ministry of Education, Culture, Sports, Science and Technology of Japan, from Project Mirai Cancer Research, and from the NOVARTIS Foundation (Japan) for the Promotion of Science (to. K. Kawada).
Acknowledgments
The authors thank Takamasa Yamamoto, Hideyo Hirai and Susumu Inamoto for data acquisition and provided facilities.
Conflicts of Interest
No potential conflict of interest exist.
Abbreviations
| CAF | cancer-associated fibroblasts |
| CRC | colorectal cancer |
| EGFR | epidermal growth factor receptor |
| FGF | fibroblast growth factor |
| G-CSF | granulocyte colony-stimulating factor |
| ICAM | intercellular adhesion molecule |
| IGF | insulin-like growth factor |
| IL | interleukin |
| MAPK | mitogen-activated protein kinase |
| MDSC | myeloid-derived suppressor cell |
| MMP | matrix metalloproteinase |
| MPE | molecular pathological epidemiology |
| NETs | neutrophil extracellular traps |
| NLR | neutrophil-to-lymphocyte ratio |
| ROS | reactive oxygen species |
| TAM | tumor-associated macrophage |
| TAN | tumor-associated neutrophil |
| TGF-β | transforming growth factor-beta |
| TIMP-1 | tissue inhibitor of metalloproteases-1 |
| TNF | tumor necrosis factor |
| VEGF | vascular endothelial growth factor |
References
- Malvezzi, M.; Bertuccio, P.; Levi, F.; Vecchia, C.L.; Negri, E. European cancer mortality predictions for the year 2014. Ann. Oncol. 2014, 25, 1650–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hori, M.; Matsuda, T.; Shibata, A.; Katanoda, K. Cancer incidence and incidence rates in Japan in 2009: A study of 32 population-based cancer registries for the Monitoring of Cancer Incidence in Japan (MCIJ) project. Jpn. J. Clin. Oncol. 2015, 45, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Ward, E.M.; Johnson, C.J.; Kathleen, A.; Ma, J.; Ryerson, A.B.; Mariotto, A.; Lake, A.J.; Wilson, R.; Sherman, R.L.; et al. Annual Report to the Nation on the Status of Cancer, 1975–2014, Featuring Survival. J. Natl. Cancer Inst. 2017, 109, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Ohhara, Y.; Fukuda, N.; Takeuchi, S.; Honma, R.; Shimizu, Y.; Kinoshita, I.; Dosaka-Akita, H. Role of targeted therapy in metastatic colorectal cancer. World J. Gastroint. Oncol. 2016, 8, 642–655. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Ward, E. International Trends in Colorectal Cancer Incidence Rates. Cancer Epidemiol. Prev. Biomark. 2009, 18, 1688–1695. [Google Scholar]
- Hackl, C.; Gerken, M.; Loss, M.; Klinkhammer-Schalke, M.; Piso, P.; Schlitt, H.J. A population-based analysis on the rate and surgical management of colorectal liver metastases in Southern Germany. Int. J. Colorectal Dis. 2011, 26, 1475–1481. [Google Scholar] [CrossRef]
- Beppu, T.; Sakamoto, Y.; Hasegawa, K.; Honda, G.; Tanaka, K.; Kotera, Y. A nomogram predicting disease-free survival in patients with colorectal liver metastases treated with hepatic resection: Multicenter data collection as a Project Study for Hepatic Surgery of the Japanese Society of Hepato-Biliary-Pancreatic Surgery. J. Hepato-Biliary-Pancreat. Sci. 2012, 19, 72–84. [Google Scholar] [CrossRef]
- Fidler, I.J. The pathogenesis of cancer metastasis: The “seed and soil” hypothesis revisited. Nat. Rev. Cancer 2003, 3, 453–458. [Google Scholar] [CrossRef]
- Anderson, R.L.; Balasas, T.; Callaghan, J.; Coombes, R.C.; Evans, J.; Hall, J.A.; Kinrade, S.; Jones, D.; Jones, P.S.; Jones, R.; et al. A framework for the development of effective anti-metastatic agents. Nat. Rev. Clin. Oncol. 2018. [Google Scholar] [CrossRef]
- Luzzi, K.J.; Macdonald, I.C.; Schmidt, E.E.; Kerkvliet, N.; Morris, V.L.; Chambers, A.F.; Groom, A.C. Multistep Nature of Metastatic Inefficiency Dormancy of Solitary Cells after Successful Extravasation and Limited Survival of Early Micrometastases. Am. J. Pathol. 1998, 153, 865–873. [Google Scholar] [CrossRef]
- Joyce, J.A.; Pollard, J.W. Microenvironmental regulation of metastasis. Nat. Rev. Cancer 2009, 9, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Orimo, A.; Gupta, P.B.; Sgroi, D.C.; Arenzana-seisdedos, F.; Delaunay, T.; Naeem, R.; Carey, V.J.; Richardson, A.L.; Weinberg, R.A. Stromal Fibroblasts Present in Invasive Human Breast Carcinomas Promote Tumor Growth and Angiogenesis through Elevated SDF-1/CXCL12 Secretion. Cell 2005, 121, 335–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karnoub, A.E.; Dash, A.B.; Vo, A.P.; Sullivan, A.; Brooks, M.W.; Bell, G.W.; Richardson, A.L.; Polyak, K.; Tubo, R.; Weinberg, R.A. Mesenchymal stem cells within tumour stroma promote breast cancer metastasis. Nature 2007, 449, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Mittal, V. The role of bone-marrow-derived cells in tumor growth, metastasis initiation and progression. Trends Mol. Med. 2009, 15, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Murdoch, C.; Muthana, M.; Coffelt, S.B.; Lewis, C.E. The role of myeloid cells in the promotion of tumour angiogenesis. Nat. Rev. Cancer 2008, 8, 618–631. [Google Scholar] [CrossRef] [PubMed]
- Gabrilovich, D.I.; Ostrand-Rosenberg, S.; Bronte, V. Coordinated regulation of myeloid cells by tumours. Nat. Rev. Immunol. 2012, 12, 253–268. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, T.; Qian, B.-Z.; Pollard, J.W. Immune cell promotion of metastasis. Nat. Rev. Immunol. 2015, 15, 73–86. [Google Scholar] [CrossRef] [Green Version]
- Engblom, C.; Pfirschke, C.; Pittet, M.J. The role of myeloid cells in cancer therapies. Nat. Rev. Cancer 2016, 16, 447–462. [Google Scholar] [CrossRef]
- Itatani, Y.; Kawada, K.; Inamoto, S.; Yamamoto, T.; Ogawa, R.; Taketo, M.M.; Sakai, Y. The Role of Chemokines in Promoting Colorectal Cancer Invasion/Metastasis. Int. J. Mol. Sci. 2016, 17, 643. [Google Scholar] [CrossRef]
- Balkwill, F. Cancer and the chemokine network. Nat. Rev. Cancer 2004, 4, 540–550. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Cassatella, M.A.; Costantini, C.; Jaillon, S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat. Rev. Immunol. 2011, 11, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eck, M.; Schmausser, B.; Scheller, K.; Brandlein, S.; Muller-Hermelink, H.K. Pleiotropic effects of CXC chemokines in gastric carcinoma: Differences in CXCL8 and CXCL1 expression between diffuse and intestinal types of gastric carcinoma. Clin. Exp. Immunol. 2003, 134, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Gijsbers, K.; Gouwy, M.; Struyf, S.; Wuyts, A.; Proost, P.; Opdenakker, G.; Penninckx, F.; Ectors, N.; Geboes, K.; Van Damme, J. GCP-2/CXCL6 synergizes with other endothelial cell-derived chemokines in neutrophil mobilization and is associated with angiogenesis in gastrointestinal tumors. Exp. Cell Res. 2005, 303, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Luan, J.; Shattuck-Brandt, R.; Haghnegahdar, H.; Owen, J.D.; Strieter, R.; Burdick, M.; Nirodi, C.; Beauchamp, D.; Johnson, K.N.; Richmond, A. Mechanism and biological significance of constitutive expression of MGSA/GRO chemokines in malignant melanoma tumor progression. J. Leukoc. Biol. 1997, 62, 588–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borregaard, N. Neutrophils, from marrow to microbes. Immunity 2010, 33, 657–670. [Google Scholar] [CrossRef]
- Nathan, C. Neutrophils and immunity: Challenges and opportunities. Nat. Rev. Immunol. 2006, 6, 173–182. [Google Scholar] [CrossRef]
- Coffelt, S.B.; Wellenstein, M.D.; de Visser, K.E. Neutrophils in cancer: Neutral no more. Nat. Rev. Cancer 2016, 16, 431–446. [Google Scholar] [CrossRef]
- Fridlender, Z.G.; Albelda, S.M. Tumor-associated neutrophils: Friend or foe? Carcinogenesis 2012, 33, 949–955. [Google Scholar] [CrossRef]
- Granot, Z.; Jablonska, J. Distinct Functions of Neutrophil in Cancer and Its Regulation. Med. Inflamm. 2015, 2015, 701067. [Google Scholar] [CrossRef] [PubMed]
- Fridlender, Z.G.; Sun, J.; Kim, S.; Kapoor, V.; Cheng, G.; Ling, L.; Worthen, G.S.; Albelda, S.M. Polarization of tumor-associated neutrophil phenotype by TGF-beta: “N1” versus “N2” TAN. Cancer Cell 2009, 16, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Shaul, M.E.; Fridlender, Z.G. Neutrophils as active regulators of the immune system in the tumor microenvironment. J. Leukoc. Biol. 2017, 102, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Houghton, A.M.; Rzymkiewicz, D.M.; Ji, H.; Gregory, A.D.; Egea, E.E.; Metz, H.E.; Stolz, D.B.; Land, S.R.; Marconcini, L.A.; Kliment, C.R.; et al. Neutrophil elastase-mediated degradation of IRS-1 accelerates lung tumor growth. Nat. Med. 2010, 16, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Kuang, D.-M.; Zhao, Q.; Wu, Y.; Peng, C.; Wang, J.; Xu, Z.; Yin, X.Y.; Zheng, L. Peritumoral neutrophils link inflammatory response to disease progression by fostering angiogenesis in hepatocellular carcinoma. J. Hepatol. 2011, 54, 948–955. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Piao, Y.; Holmes, L.; Fuller, G.N.; Henry, V.; Tiao, N.; de Groot, J.F. Neutrophils promote the malignant glioma phenotype through S100A4. Clin. Cancer Res. 2014, 20, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Wislez, M.; Rabbe, N.; Marchal, J.; Milleron, B.; Crestani, B.; Mayaud, C.; Antoine, M.; Soler, P.; Cadranel, J. Hepatocyte growth factor production by neutrophils infiltrating bronchioloalveolar subtype pulmonary adenocarcinoma: Role in tumor progression and death. Cancer Res. 2003, 63, 1405–1412. [Google Scholar]
- Andzinski, L.; Kasnitz, N.; Stahnke, S.; Wu, C.-F.; Gereke, M.; von Kockritz-Blickwede, M.; Schilling, S.; Brandau, S.; Weiss, S.; Jablonska, J. Type I IFNs induce anti-tumor polarization of tumor associated neutrophils in mice and human. Int. J. Cancer 2016, 138, 1982–1993. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Yipp, B.G.; Kubes, P. NETosis: How vital is it? Blood 2013, 122, 2784–2794. [Google Scholar] [CrossRef] [PubMed]
- Jaillon, S.; Peri, G.; Delneste, Y.; Fremaux, I.; Doni, A.; Moalli, F.; Garlanda, C.; Romani, L.; Gascan, H.; Bellocchio, S.; et al. The humoral pattern recognition receptor PTX3 is stored in neutrophil granules and localizes in extracellular traps. J. Exp. Med. 2007, 204, 793–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.H.; Fraser, I.P.; Fukase, K.; Kusumoto, S.; Fujimoto, Y.; Stahl, G.L.; Ezekowitz, R.A. Human peptidoglycan recognition protein S is an effector of neutrophil-mediated innate immunity. Blood 2005, 106, 2551–2558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dziarski, R.; Platt, K.A.; Gelius, E.; Steiner, H.; Gupta, D. Defect in neutrophil killing and increased susceptibility to infection with nonpathogenic gram-positive bacteria in peptidoglycan recognition protein-S (PGRP-S)-deficient mice. Blood 2003, 102, 689–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urban, C.F.; Ermert, D.; Schmid, M.; Abu-Abed, U.; Goosmann, C.; Nacken, W.; Brinkmann, V.; Jungblut, P.R.; Zychlinsky, A. Neutrophil extracellular traps co.ntain calprotectin, a cytosolic protein complex involved in host defense against Candida albicans. PLoS Pathog. 2009, 5, e1000639. [Google Scholar] [CrossRef] [PubMed]
- Cools-Lartigue, J.; Spicer, J.; McDonald, B.; Gowing, S.; Chow, S.; Giannias, B.; Bourdeau, F.; Kubes, P.; Ferri, L. Neutrophil extracellular traps sequester circulating tumor cells and promote metastasis. J. Clin. Investig. 2013, 123, 3446–3458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demers, M.; Wagner, D.D. NETosis: A new factor in tumor progression and cancer-associated thrombosis. Semin. Thromb. Hemost. 2014, 40, 277–283. [Google Scholar] [CrossRef]
- Tohme, S.; Yazdani, H.O.; Al-Khafaji, A.B.; Chidi, A.P.; Loughran, P.; Mowen, K.; Wang, Y.; Simmons, R.L.; Huang, H.; Tsung, A. Neutrophil Extracellular Traps Promote the Development and Progression of Liver Metastases after Surgical Stress. Cancer Res. 2016, 76, 1367–1380. [Google Scholar] [CrossRef] [Green Version]
- Najmeh, S.; Cools-Lartigue, J.; Rayes, R.F.; Gowing, S.; Vourtzoumis, P.; Bourdeau, F.; Giannias, B.; Berube, J.; Rousseau, S.; Ferri, L.E.; et al. Neutrophil extracellular traps sequester circulating tumor cells via beta1-integrin mediated interactions. Int. J. Cancer 2017, 140, 2321–2330. [Google Scholar] [CrossRef]
- Coussens, L.M.; Tinkle, C.L.; Hanahan, D.; Werb, Z. MMP-9 supplied by bone marrow-derived cells contributes to skin carcinogenesis. Cell 2000, 103, 481–490. [Google Scholar] [CrossRef]
- Bergers, G.; Brekken, R.; McMahon, G.; Vu, T.H.; Itoh, T.; Tamaki, K.; Tanzawa, K.; Thorpe, P.; Itohara, S.; Werb, Z.; et al. Matrix metalloproteinase-9 triggers the angiogenic switch during carcinogenesis. Nat. Cell Biol. 2000, 2, 737–744. [Google Scholar] [CrossRef]
- Nozawa, H.; Chiu, C.; Hanahan, D. Infiltrating neutrophils mediate the initial angiogenic switch in a mouse model of multistage carcinogenesis. Proc. Natl. Acad. Sci. USA 2006, 103, 12493–12498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shojaei, F.; Wu, X.; Zhong, C.; Yu, L.; Liang, X.H.; Yao, J.; Blanchard, D.; Bais, C.; Peale, F.V.; van Bruggen, N.; et al. Bv8 regulates myeloid-cell-dependent tumour angiogenesis. Nature 2007, 450, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Zhuang, G.; Yu, L.; Meng, G.; Ferrara, N. Induction of Bv8 expression by granulocyte colony-stimulating factor in CD11b+Gr1+ cells: Key role of Stat3 signaling. J. Biol. Chem. 2012, 287, 19574–19584. [Google Scholar] [CrossRef] [PubMed]
- Kowanetz, M.; Wu, X.; Lee, J.; Tan, M.; Hagenbeek, T.; Qu, X.; Yu, L.; Ross, J.; Korsisaari, N.; Cao, T.; et al. Granulocyte-colony stimulating factor promotes lung metastasis through mobilization of Ly6G+Ly6C+ granulocytes. Proc. Natl. Acad. Sci. USA 2010, 107, 21248–21255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shojaei, F.; Wu, X.; Qu, X.; Kowanetz, M.; Yu, L.; Tan, M.; Meng, Y.G.; Ferrara, N. G-CSF-initiated myeloid cell mobilization and angiogenesis mediate tumor refractoriness to anti-VEGF therapy in mouse models. Proc. Natl. Acad. Sci. USA 2009, 106, 6742–6747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eash, K.J.; Greenbaum, A.M.; Gopalan, P.K.; Link, D.C. CXCR2 and CXCR4 antagonistically regulate neutrophil trafficking from murine bone marrow. J. Clin. Investig. 2010, 120, 2423–2431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Highfill, S.L.; Cui, Y.; Giles, A.J.; Smith, J.P.; Zhang, H.; Morse, E.; Kaplan, R.N.; Mackall, C.L. Disruption of CXCR2-mediated MDSC tumor trafficking enhances anti-PD1 efficacy. Sci. Transl. Med. 2014, 6, 237ra67. [Google Scholar] [CrossRef]
- Steele, C.W.; Karim, S.A.; Leach, J.D.G.; Bailey, P.; Upstill-Goddard, R.; Rishi, L.; Foth, M.; Bryson, S.; McDaid, K.; Wilson, Z.; et al. CXCR2 Inhibition Profoundly Suppresses Metastases and Augments Immunotherapy in Pancreatic Ductal Adenocarcinoma. Cancer Cell 2016, 29, 832–845. [Google Scholar] [CrossRef] [Green Version]
- Jamieson, T.; Clarke, M.; Steele, C.W.; Samuel, M.S.; Neumann, J.; Jung, A.; Huels, D.; Olson, M.F.; Das, S.; Nibbs, R.J.; et al. Inhibition of CXCR2 profoundly suppresses inflammation-driven and spontaneous tumorigenesis. J. Clin. Investig. 2012, 122, 3127–3144. [Google Scholar] [CrossRef] [Green Version]
- Katoh, H.; Wang, D.; Daikoku, T.; Sun, H.; Dey, S.K.; Dubois, R.N. CXCR2-expressing myeloid-derived suppressor cells are essential to promote colitis-associated tumorigenesis. Cancer Cell 2013, 24, 631–644. [Google Scholar] [CrossRef]
- Nywening, T.M.; Belt, B.A.; Cullinan, D.R.; Panni, R.Z.; Han, B.J.; Sanford, D.E.; Jacobs, R.C.; Ye, J.; Patel, A.A.; Gillanders, W.E.; et al. Targeting both tumour-associated CXCR2(+) neutrophils and CCR2(+) macrophages disrupts myeloid recruitment and improves chemotherapeutic responses in pancreatic ductal adenocarcinoma. Gut 2018, 67, 1112–1123. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, R.; Yamamoto, T.; Hirai, H.; Hanada, K.; Kiyasu, Y.; Nishikawa, G.; Mizuno, R.; Inamoto, S.; Itatani, Y.; Sakai, Y.; et al. Loss of SMAD4 promotes colorectal cancer progression by recruiting tumor-associated neutrophils via CXCL1/8-CXCR2 axis. Clin. Cancer Res. (under review).
- Donskov, F. Immunomonitoring and prognostic relevance of neutrophils in clinical trials. Semin. Cancer Biol. 2013, 23, 200–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Templeton, A.J.; McNamara, M.G.; Seruga, B.; Vera-Badillo, F.E.; Aneja, P.; Ocana, A.; Leibowitz-Amit, R.; Sonpavde, G.; Knox, J.J.; Tran, B.; et al. Prognostic role of neutrophil-to-lymphocyte ratio in solid tumors: A systematic review and meta-analysis. J. Natl. Cancer Inst. 2014, 106, dju124. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, G.J.K.; Charles, K.A.; Roxburgh, C.S.D.; Horgan, P.G.; McMillan, D.C.; Clarke, S.J. The systemic inflammation-based neutrophil-lymphocyte ratio: Experience in patients with cancer. Crit. Rev. Oncol. Hematol. 2013, 88, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhao, R.; Cui, Y.; Zhou, Y.; Wu, X. The dynamic change of neutrophil to lymphocyte ratio can predict clinical outcome in stage I-III colon cancer. Sci. Rep. 2018, 8, 9453. [Google Scholar] [CrossRef]
- Borazan, E.; Balik, A.A.; Bozdag, Z.; Arik, M.K.; Aytekin, A.; Yilmaz, L.; Elqi, M.; Baskonus, I. Assessment of the relationship between neutrophil lymphocyte ratio and prognostic factors in non-metastatic colorectal cancer. Turk. J. Surg. 2017, 33, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Rashtak, S.; Ruan, X.; Druliner, B.R.; Liu, H.; Therneau, T.; Mouchli, M.; Boardman, L.A. Peripheral Neutrophil to Lymphocyte Ratio Improves Prognostication in Colon Cancer. Clin. Colorectal Cancer 2017, 16, 115–123. [Google Scholar] [CrossRef]
- Halazun, K.J.; Aldoori, A.; Malik, H.Z.; Al-Mukhtar, A.; Prasad, K.R.; Toogood, G.J.; Lodge, J.P. Elevated preoperative neutrophil to lymphocyte ratio predicts survival following hepatic resection for colorectal liver metastases. Eur. J. Surg. Oncol. 2008, 34, 55–60. [Google Scholar] [CrossRef]
- Dell’Aquila, E.; Cremolini, C.; Zeppola, T.; Lonardi, S.; Bergamo, F.; Masi, G.; Stellato, M.; Marmorino, F.; Schirripa, M.; Urbano, F.; et al. Prognostic and predictive role of neutrophil/lymphocytes ratio in metastatic colorectal cancer: A retrospective analysis of the TRIBE study by GONO. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2018, 29, 924–930. [Google Scholar] [CrossRef]
- Rao, H.-L.; Chen, J.-W.; Li, M.; Xiao, Y.-B.; Fu, J.; Zeng, Y.-X.; Cai, M.Y.; Xie, D. Increased intratumoral neutrophil in colorectal carcinomas correlates closely with malignant phenotype and predicts patients’ adverse prognosis. PLoS ONE 2012, 7, e30806. [Google Scholar] [CrossRef] [PubMed]
- Berry, R.S.; Xiong, M.-J.; Greenbaum, A.; Mortaji, P.; Nofchissey, R.A.; Schultz, F.; Martinez, C.; Luo, L.; Morris, K.T.; Hanson, J.A. High levels of tumor-associated neutrophils are associated with improved overall survival in patients with stage II colorectal cancer. PLoS ONE 2017, 12, e0188799. [Google Scholar] [CrossRef] [PubMed]
- Itatani, Y.; Kawada, K.; Fujishita, T.; Kakizaki, F.; Hirai, H.; Matsumoto, T.; Iwamoto, M.; Inamoto, S.; Hatano, E.; Hasegawa, S.; et al. Loss of SMAD4 from colorectal cancer cells promotes CCL15 expression to recruit CCR1+ myeloid cells and facilitate liver metastasis. Gastroenterology 2013, 145, 1064–1075. [Google Scholar] [CrossRef] [PubMed]
- Inamoto, S.; Itatani, Y.; Yamamoto, T.; Minamiguchi, S.; Hirai, H.; Iwamoto, M.; Hasegawa, S.; Taketo, M.M.; Sakai, Y.; Kawada, K. Loss of SMAD4 Promotes Colorectal Cancer Progression by Accumulation of Myeloid-Derived Suppressor Cells through the CCL15-CCR1 Chemokine Axis. Clin. Cancer Res. 2016, 22, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Kawada, K.; Itatani, Y.; Inamoto, S.; Okamura, R.; Iwamoto, M.; Miyamoto, E.; Chen-Yoshikawa, T.F.; Hirai, H.; Hasegawa, S.; et al. Loss of SMAD4 Promotes Lung Metastasis of Colorectal Cancer by Accumulation of CCR1+ Tumor-Associated Neutrophils through CCL15-CCR1 Axis. Clin. Cancer Res. 2017, 23, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Bronte, V.; Brandau, S.; Chen, S.-H.; Colombo, M.P.; Frey, A.B.; Greten, T.F.; Mandruzzato, S.; Murray, P.J.; Ochoa, A.; OStrand-Rosenberg, S.; et al. Recommendations for myeloid-derived suppressor cell nomenclature and characterization standards. Nat. Commun. 2016, 7, 12150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zilio, S.; Serafini, P. Neutrophils and Granulocytic MDSC: The Janus God of Cancer Immunotherapy. Vaccines 2016, 4, 31. [Google Scholar] [CrossRef]
- Hirai, H.; Fujishita, T.; Kurimoto, K.; Miyachi, H.; Kitano, S.; Inamoto, S.; Itatani, Y.; Saitou, M.; Maekawa, T.; Taketo, M.M. CCR1-mediated accumulation of myeloid cells in the liver microenvironment promoting mouse colon cancer metastasis. Clin. Exp. Metastasis 2014, 31, 977–989. [Google Scholar] [CrossRef] [Green Version]
- Rodero, M.P.; Auvynet, C.; Poupel, L.; Combadiere, B.; Combadiere, C. Control of both myeloid cell infiltration and angiogenesis by CCR1 promotes liver cancer metastasis development in mice. Neoplasia 2013, 15, 641–648. [Google Scholar] [CrossRef]
- Kiyasu, Y.; Kyoto University, Kyoto, Japan. Personal communication, 2018.
- Kumar, A.; Cherukumilli, M.; Mahmoudpour, S.H.; Brand, K.; Bandapalli, O.R. ShRNA-mediated knock-down of CXCL8 inhibits tumor growth in colorectal liver metastasis. Biochem. Biophys. Res. Commun. 2018, 500, 731–737. [Google Scholar] [CrossRef]
- Yamamoto, M.; Kikuchi, H.; Ohta, M.; Kawabata, T.; Hiramatsu, Y.; Kondo, K.; Baba, M.; Kamiya, K.; Tanaka, T.; Kitagawa, M.; et al. TSU68 prevents liver metastasis of colon cancer xenografts by modulating the premetastatic niche. Cancer Res. 2008, 68, 9754–9762. [Google Scholar] [CrossRef] [PubMed]
- Varney, M.L.; Singh, S.; Li, A.; Mayer-Ezell, R.; Bond, R.; Singh, R.K. Small molecule antagonists for CXCR2 and CXCR1 inhibit human colon cancer liver metastases. Cancer Lett. 2011, 300, 180–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon-Weeks, A.N.; Lim, S.Y.; Yuzhalin, A.E.; Jones, K.; Markelc, B.; Kim, K.J.; Buzzelli, J.N.; Fokas, E.; Cao, Y.; Smart, S.; et al. Neutrophils promote hepatic metastasis growth through fibroblast growth factor 2-dependent angiogenesis in mice. Hepatology 2017, 65, 1920–1935. [Google Scholar] [CrossRef] [PubMed]
- Rayes, R.F.; Milette, S.; Fernandez, M.C.; Ham, B.; Wang, N.; Bourdeau, F.; Perrino, S.; Yakar, S.; Brodt, P. Loss of neutrophil polarization in colon carcinoma liver metastases of mice with an inducible, liver-specific IGF-I deficiency. Oncotarget 2018, 9, 15691–15704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seubert, B.; Grunwald, B.; Kobuch, J.; Cui, H.; Schelter, F.; Schaten, S.; Siveke, J.T.; Lim, N.H.; Nagase, H.; Simonavicius, N.; et al. Tissue inhibitor of metalloproteinases (TIMP)-1 creates a premetastatic niche in the liver through SDF-1/CXCR4-dependent neutrophil recruitment in mice. Hepatology 2015, 61, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Aoki, T.; Tsuruyama, T.; Narumiya, S. Definition of Prostaglandin E2-EP2 Signals in the Colon Tumor Microenvironment That Amplify Inflammation and Tumor Growth. Cancer Res. 2015, 75, 2822–2832. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.K.; Eckmann, L.; Panja, A.; Kagnoff, M.F. Differential and regulated expression of C-X-C, C-C, and C-chemokines by human colon epithelial cells. Gastroenterology 1997, 113, 1214–1223. [Google Scholar] [CrossRef]
- Mizukami, Y.; Jo, W.-S.; Duerr, E.-M.; Gala, M.; Li, J.; Zhang, X.; Zimmer, M.A.; Iliopoulos, O.; Zukerberg, L.R.; Kohgo, Y.; et al. Induction of interleukin-8 preserves the angiogenic response in HIF-1alpha-deficient colon cancer cells. Nat. Med. 2005, 11, 992–997. [Google Scholar] [CrossRef]
- Ning, Y.; Manegold, P.C.; Hong, Y.K.; Zhang, W.; Pohl, A.; Lurje, G.; Winder, T.; Yang, D.; LaBonte, M.J.; Wilson, P.M.; et al. Interleukin-8 is associated with proliferation, migration, angiogenesis and chemosensitivity in vitro and in vivo in colon cancer cell line models. Int. J. Cancer 2011, 128, 2038–2049. [Google Scholar] [CrossRef]
- Lee, Y.S.; Choi, I.; Ning, Y.; Kim, N.Y.; Khatchadourian, V.; Yang, D.; Chung, H.K.; Choi, D.; LaBonte, M.J.; Ladner, R.D.; et al. Interleukin-8 and its receptor CXCR2 in the tumour microenvironment promote colon cancer growth, progression and metastasis. Br. J. Cancer 2012, 106, 1833–1841. [Google Scholar] [CrossRef] [Green Version]
- Ning, Y.; Labonte, M.J.; Zhang, W.; Bohanes, P.O.; Gerger, A.; Yang, D.; Benhaim, L.; Paez, D.; Rosenberg, D.O.; Nagulapalli Venkata, K.C.; et al. The CXCR2 antagonist, SCH-527123, shows antitumor activity and sensitizes cells to oxaliplatin in preclinical colon cancer models. Mol. Cancer Ther. 2012, 11, 1353–1364. [Google Scholar] [CrossRef] [PubMed]
- Asfaha, S.; Dubeykovskiy, A.; Tomita, H.; Yang, X.; Stokes, S.; Shibata, W.; Friedman, R.A.; Ariyama, H.; Dubeykovskaya, Z.A.; Muthupalani, S.; et al. Mice that express human interleukin-8 have increased mobilization of immature myeloid cells, which exacerbates inflammation and accelerates colon carcinogenesis. Gastroenterology 2013, 144, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Glasser, L.; Fiederlein, R.L. The effect of various cell separation procedures on assays of neutrophil function. A critical appraisal. Am. J. Clin. Pathol. 1990, 93, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Kostic, A.D.; Chun, E.; Meyerson, M.; Garrett, W.S. Microbes and inflammation in colorectal cancer. Cancer Immunol. Res. 2013, 1, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Ogino, S.; Chan, A.T.; Fuchs, C.S.; Giovannucci, E. Molecular pathological epidemiology of colorectal neoplasia: An emerging transdisciplinary and interdisciplinary field. Gut 2011, 60, 394–411. [Google Scholar] [CrossRef] [PubMed]
- Ogino, S.; Nowak, J.A.; Hamada, T.; Phipps, A.I.; Peters, U.; Milner, D.A., Jr.; Giovannucci, E.L.; Nishihara, R.; Giannakis, M.; Garrett, W.S.; et al. Integrative analysis of exogenous, endogenous, tumour and immune factors for precision medicine. Gut 2018, 67, 1168–1180. [Google Scholar] [CrossRef] [PubMed]
- Bierie, B.; Moses, H.L. Tumour microenvironment: TGFbeta: The molecular Jekyll and Hyde of cancer. Nat. Rev. Cancer 2006, 6, 506–520. [Google Scholar] [CrossRef] [PubMed]
- Shaul, M.E.; Fridlender, Z.G. Cancer-related circulating and tumor-associated neutrophils - subtypes, sources and function. FEBS J. 2018, 285, 4316–4342. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Carroll, K.D.; Policarpio, D.; Osborn, C.; Gregory, M.; Bassi, R.; Jimenez, X.; Prewett, M.; Liebisch, G.; Persaud, K.; et al. Anti-transforming growth factor beta receptor II antibody has therapeutic efficacy against primary tumor growth and metastasis through multieffects on cancer, stroma, and immune cells. Clin. Cancer Res. 2010, 16, 1191–1205. [Google Scholar] [CrossRef]
- Akhurst, R.J. Targeting TGF-beta Signaling for Therapeutic Gain. Cold Spring Harb. Perspect. Biol. 2017, 9, a022301. [Google Scholar] [CrossRef]
Figure 1.
Overview of the process of liver metastasis.


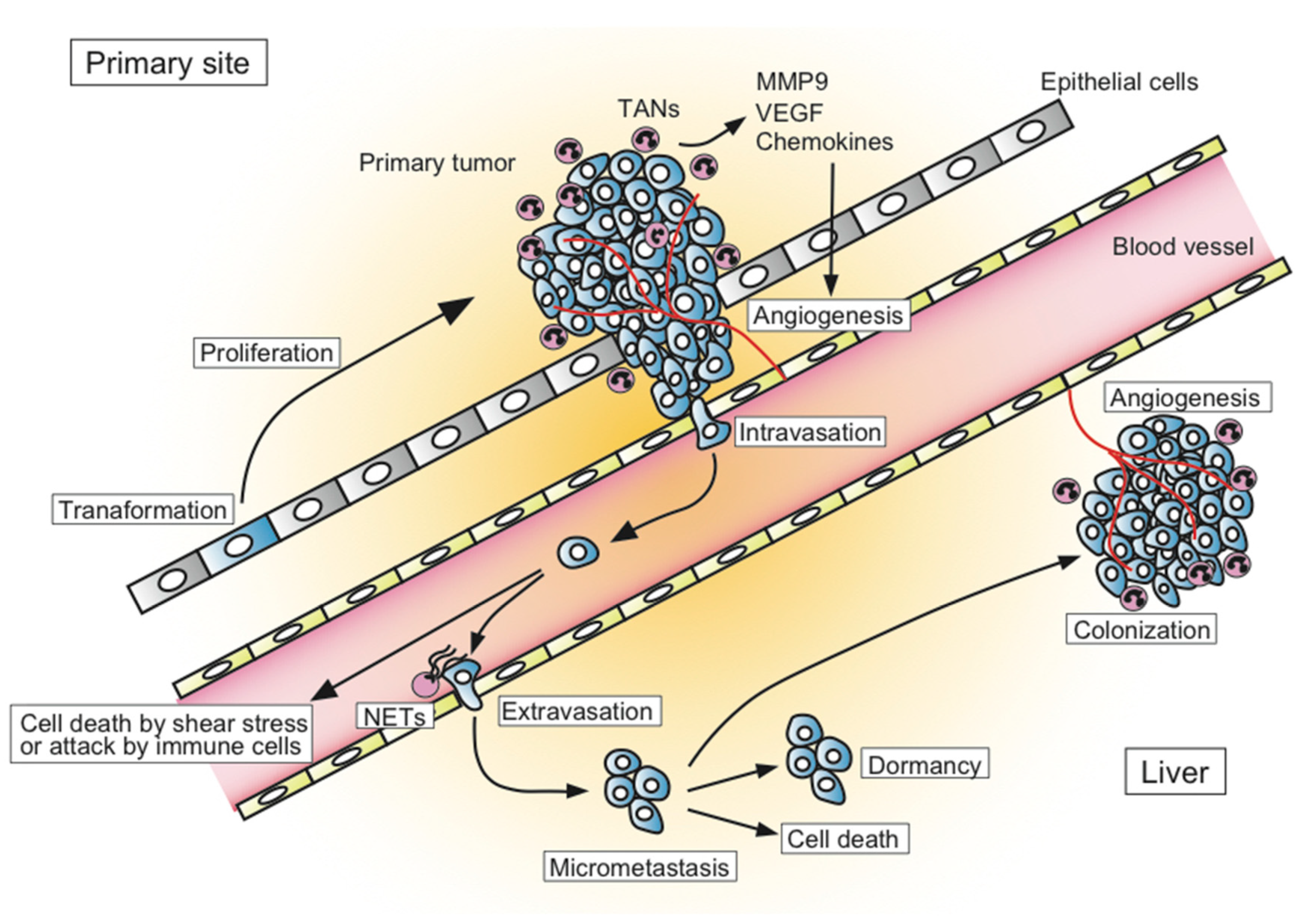{kind=link}
Table 1.
TANs in animal models for CRC metastasis.
| Authors | Reference | Animal | Cell | Molecules | Effect |
|---|---|---|---|---|---|
| Hirai et al. | [78] | Mouse | CRC cells | CCL-9 | CCL-9 in CRC cells recruit CCR1+ neutrophils which produce MMP9 for cancer expansion |
| Rodero et al. | [79] | Mouse | Hematopoietic/nonhematopoietic cells | CCR1 | CCR1 expression by both hematopoietic and non-hematopoietic cells favors tumor aggressiveness |
| Kiyasu, Y. | [80] | Mouse | Myeloid cells | CCR1 | CCR1-knockout in myeloid cells suppress CRC liver metastasis |
| Kumar et al. | [81] | Mouse | CRC cells | CXCL8 | CXCL8 promotes neutrophil recruitment, metastasis, angiogenesis and invasion |
| Yamamoto et al. | [82] | Mouse | CXCL1/CXCR2 | CXCL1/CXCR2 axis is important in cancer metastasis | |
| Varney et al. | [83] | Mouse | Systemic | CXCR1/CXCR2 | Systemic inhibition of CXCR1/CXCR2 induced apoptosis and inhibited angiogenesis in the liver metastasis |
| Gordon-Weeks et al. | [84] | Mouse | TANs | FGF2 | FGF2 in TANs induce polarization of neutrophils |
| Rayes et al. | [85] | Mouse | Liver | IGF-1 | Sustained IGF-1 deficiency in liver alters the neutrophil phenotypes |
| Seubert et al. | [86] | Mouse | Systemic | TIMP-1 | Systemic TIMP-1 expression promotes neutrophil recruitment through the increase of hepatic SDF-1 and increase the liver susceptibility |
| Ma et al. | [87] | Mouse | TANs | EP2 | EP2 signaling in TANs promotes tumor growth through TNF-β, IL-6, CXCL1, COX-2, and Wnt5A |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mizuno, R.; Kawada, K.; Itatani, Y.; Ogawa, R.; Kiyasu, Y.; Sakai, Y. The Role of Tumor-Associated Neutrophils in Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 529. https://doi.org/10.3390/ijms20030529
AMA Style
Mizuno R, Kawada K, Itatani Y, Ogawa R, Kiyasu Y, Sakai Y. The Role of Tumor-Associated Neutrophils in Colorectal Cancer. International Journal of Molecular Sciences. 2019; 20(3):529. https://doi.org/10.3390/ijms20030529
Chicago/Turabian StyleMizuno, Rei, Kenji Kawada, Yoshiro Itatani, Ryotaro Ogawa, Yoshiyuki Kiyasu, and Yoshiharu Sakai. 2019. "The Role of Tumor-Associated Neutrophils in Colorectal Cancer" International Journal of Molecular Sciences 20, no. 3: 529. https://doi.org/10.3390/ijms20030529
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.