Assessment of miR-98-5p, miR-152-3p, miR-326 and miR-4289 Expression as Biomarker for Prostate Cancer Diagnosis

 ,
,
Abstract
:1. Introduction
2. Results
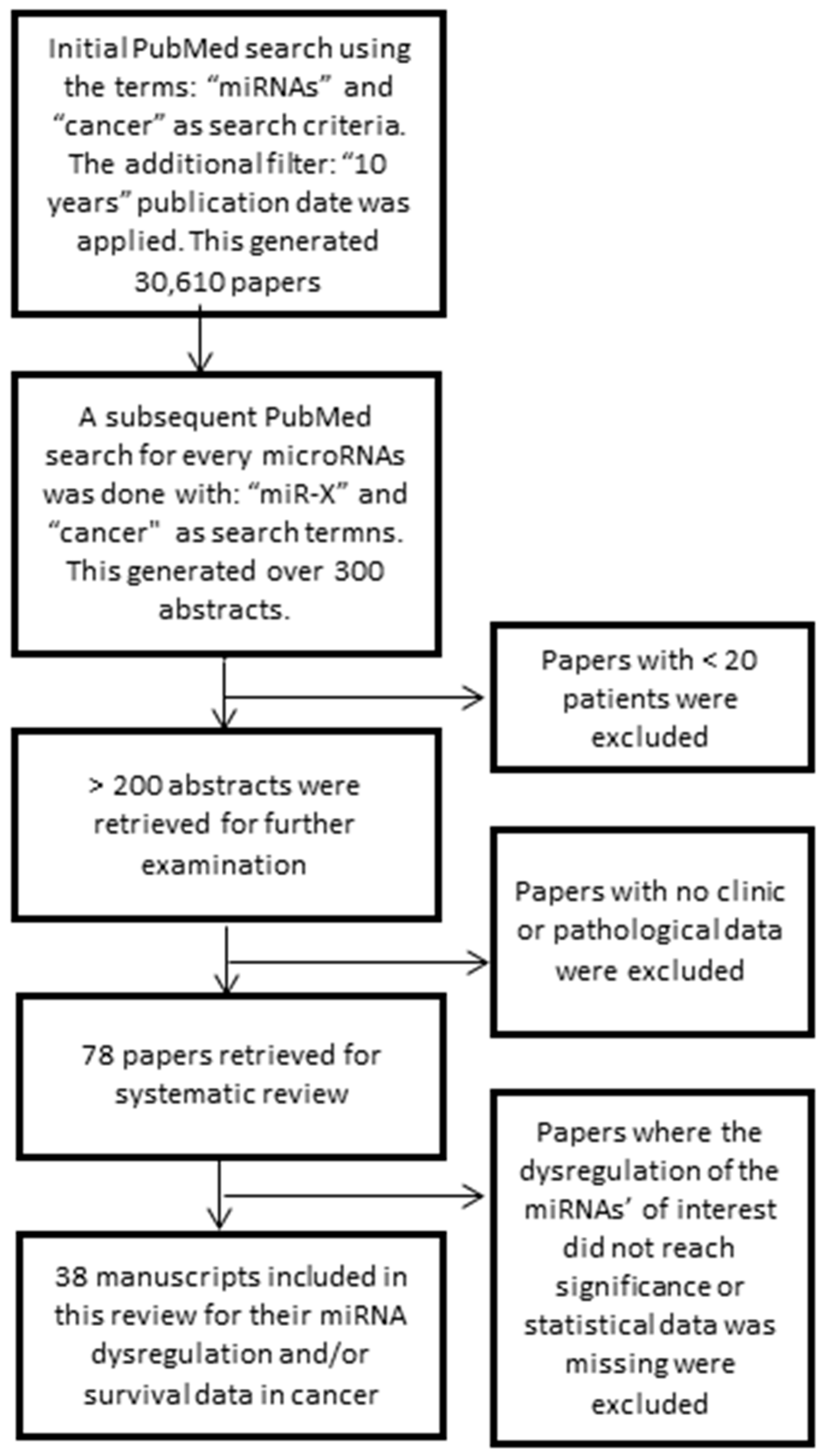
2.1. Literature Retrieval
2.2. Dysregulation of miR-98-5p, miR-152-3p, miR-326 and miR-4289 in Cancer Patient Samples
2.2.1. Dysregulation of miR-98-5p in Cancer
2.2.2. Dysregulation of miR-152-3p in Cancer
2.2.3. Dysregulation of miR-326 in Cancer
2.2.4. Dysregulation of miR-4289 in Cancer
2.3. Differential Expression of the miRNA Signature in Cancerous Samples when Compared to Non-Cancerous Samples from Microarray Dataset
2.4. Functional Role of the miRNA Signature in Cancer
3. Discussion
4. Materials and Methods
4.1. Research Data Extraction Strategy: Inclusion and Exclusion Criteria
4.2. Differential Expression of the miRNA Signature in Cancer: High-Throughput Data
4.3. Dysregulation of miR-98-5p, miR-152-3p, miR-326 and miR-4289 Associated with Cancer Prognosis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Stamey, T.A.; Yang, N.; Hay, A.R.; McNeal, J.E.; Freiha, F.S.; Redwine, E. Prostate-specific antigen as a serum marker for adenocarcinoma of the prostate. N. Engl. J. Med. 1987, 317, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Diamandis, E.P. Prostate-specific Antigen: Its Usefulness in Clinical Medicine. Trends Endocrinol. Metab. 1998, 9, 310–316. [Google Scholar] [CrossRef]
- Adhyam, M.; Gupta, A.K. A Review on the Clinical Utility of PSA in Cancer Prostate. Indian J. Surg. Oncol. 2012, 3, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Bokhorst, L.P.; Bangma, C.H.; van Leenders, G.J.; Lous, J.J.; Moss, S.M.; Schroder, F.H.; Roobol, M.J. Prostate-specific antigen-based prostate cancer screening: Reduction of prostate cancer mortality after correction for nonattendance and contamination in the Rotterdam section of the European Randomized Study of Screening for Prostate Cancer. Eur. Urol. 2014, 65, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Carter, H.; Albersten, P.; Barry, M.J.; Etzioni, R.; Freedland, S.; Greene, K.; Holmberg, L.; Kantoff, P.; Koneti, B.; Murad, M.; et al. Early Detection of Prostate Cancer. J. Urol. 2015. [Google Scholar] [CrossRef]
- U.S. Preventative Services Task Force. Prostate Cancer Screening Draft Recommendations; USPSTF: Washington, DC, USA, 2017. [Google Scholar]
- Cancer Council Australia. Prostate Cancer; Cancer Council Australia: Sydney, Australia, 2017. [Google Scholar]
- Fradet, Y.; Klotz, L.; Trachtenberg, J.; Zlotta, A. The burden of prostate cancer in Canada. Can. Urol. Assoc. J. 2009, 3 (Suppl. 2), S92–S100. [Google Scholar] [CrossRef] [PubMed]
- Grover, S.A.; Coupal, L.; Zowall, H.; Rajan, R.; Trachtenberg, J.; Elhilali, M.; Chetner, M.; Goldenberg, L. The economic burden of prostate cancer in Canada: Forecasts from the Montreal Prostate Cancer Model. Can. Med. Assoc. J. 2000, 162, 987–992. [Google Scholar]
- Walter, B.A.; Valera, V.A.; Pinto, P.A.; Merino, M.J. Comprehensive microRNA Profiling of Prostate Cancer. J. Cancer 2013, 4, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef]
- Macfarlane, L.A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [PubMed]
- Lovat, F.; Valeri, N.; Croce, C.M. MicroRNAs in the pathogenesis of cancer. Semin. Oncol. 2011, 38, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Mazan-Mamczarz, K.; Gartenhaus, R.B. Role of microRNA deregulation in the pathogenesis of diffuse large B-cell lymphoma (DLBCL). Leuk Res. 2013, 37, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, M.; Li, Y.; Ye, S.; Ma, J.; Lu, L.; Lv, W.; Chang, G.; Li, X.; Li, Q.; Wang, S.; et al. MicroRNA profiling implies new markers of chemoresistance of triple-negative breast cancer. PLoS ONE 2014, 9, e96228. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Wu, W.K.; Wu, C.W.; Sung, J.J.; Yu, J.; Ng, S.S. MicroRNA dysregulation in colorectal cancer: A clinical perspective. Br. J. Cancer 2011, 104, 893–898. [Google Scholar] [CrossRef] [PubMed]
- Maugeri-Sacca, M.; Coppola, V.; Bonci, D.; De Maria, R. MicroRNAs and prostate cancer: From preclinical research to translational oncology. Cancer J. 2012, 18, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Tumilson, C.A.; Lea, R.W.; Alder, J.E.; Shaw, L. Circulating microRNA biomarkers for glioma and predicting response to therapy. Mol. Neurobiol. 2014, 50, 545–558. [Google Scholar] [CrossRef] [PubMed]
- Matin, F.; Batra, J. A microRNA molecular signature of aggressive prostate cancer. Transl. Cancer Res. 2017, 6, S12–S14. [Google Scholar] [CrossRef]
- Matin, F.; Jeet, V.; Clements, J.A.; Yousef, G.M.; Batra, J. MicroRNA Theranostics in Prostate Cancer Precision Medicine. Clin. Chem. 2016, 62, 1318–1333. [Google Scholar] [CrossRef]
- Mengual, L.; Lozano, J.J.; Ingelmo-Torres, M.; Gazquez, C.; Ribal, M.J.; Alcaraz, A. Using microRNA profiling in urine samples to develop a non-invasive test for bladder cancer. Int. J. Cancer 2013, 133, 2631–2641. [Google Scholar]
- Xing, L.; Todd, N.W.; Yu, L.; Fang, H.; Jiang, F. Early detection of squamous cell lung cancer in sputum by a panel of microRNA markers. Mod. Pathol. 2010, 23, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Ferracin, M.; Pedriali, M.; Veronese, A.; Zagatti, B.; Gafa, R.; Magri, E.; Lunardi, M.; Munerato, G.; Querzoli, G.; Maestri, I.; et al. MicroRNA profiling for the identification of cancers with unknown primary tissue-of-origin. J. Pathol. 2011, 225, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Youssef, Y.M.; White, N.M.; Grigull, J.; Krizova, A.; Samy, C.; Mejia-Guerrero, S.; Evans, A.; Yousef, G.M. Accurate molecular classification of kidney cancer subtypes using microRNA signature. Eur. Urol. 2011, 59, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Matin, F.; Jeet, V.; Moya, L.; Selth, L.A.; Chambers, S.; Yeadon, T.; Saunders, P.; Eckert, A.; Heathcote, P.; Wood, G.; et al. A Plasma Biomarker Panel of Four MicroRNAs for the Diagnosis of Prostate Cancer. Sci. Rep. 2018, 8, 6653. [Google Scholar] [CrossRef] [PubMed]
- Thompson, I.M.; Ankerst, D.P.; Chi, C.; Lucia, M.S.; Goodman, P.J.; Crowley, J.J.; Parnes, H.L.; Coltman, C.A. Jr. Operating characteristics of prostate-specific antigen in men with an initial PSA level of 3.0 ng/mL or lower. JAMA 2005, 294, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Lilja, H.; Ulmert, D.; Vickers, A.J. Prostate-specific antigen and prostate cancer: Prediction, detection and monitoring. Nat. Rev. Cancer 2008, 8, 268. [Google Scholar] [CrossRef] [PubMed]
- Pashaei, E.; Pashaei, E.; Ahmady, M.; Ozen, M.; Aydin, N. Meta-analysis of miRNA expression profiles for prostate cancer recurrence following radical prostatectomy. PLoS ONE 2017, 12, e0179543. [Google Scholar] [CrossRef]
- Zhou, W.; Zou, B.; Liu, L.; Cui, K.; Gao, J.; Yuan, S.; Cong, N. MicroRNA-98 acts as a tumor suppressor in hepatocellular carcinoma via targeting SALL4. Oncotarget 2016, 7, 74059–74073. [Google Scholar] [CrossRef]
- Wang, C.Y.; Zhang, J.J.; Hua, L.; Yao, K.H.; Chen, J.T.; Ren, X.Q. MicroRNA-98 suppresses cell proliferation, migration and invasion by targeting collagen triple helix repeat containing 1 in hepatocellular carcinoma. Mol. Med. Rep. 2016, 13, 2639–2644. [Google Scholar] [CrossRef]
- Huang, S.D.; Yuan, Y.; Zhuang, C.W.; Li, B.L.; Gong, D.J.; Wang, S.G.; Zeng, Z.Y.; Cheng, H.Z. MicroRNA-98 and microRNA-214 post-transcriptionally regulate enhancer of zeste homolog 2 and inhibit migration and invasion in human esophageal squamous cell carcinoma. Mol. Cancer 2012, 11, 51. [Google Scholar] [CrossRef]
- Chen, Z.; Cheng, Q.; Ma, Z.; Xi, H.; Peng, R.; Jiang, B. Overexpression of RKIP Inhibits Cell Invasion in Glioma Cell Lines through Upregulation of miR-98. BioMed. Res. Int. 2013, 2013, 695179. [Google Scholar] [CrossRef] [PubMed]
- Ni, R.; Huang, Y.; Wang, J. miR-98 targets ITGB3 to inhibit proliferation, migration, and invasion of non-small-cell lung cancer. OncoTargets Ther. 2015, 8, 2689–2697. [Google Scholar]
- Wang, K.; Dong, L.; Fang, Q.; Xia, H.; Hou, X. Low serum miR-98 as an unfavorable prognostic biomarker in patients with non-small cell lung cancer. Cancer Biomark. 2017, 20, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, X.J.; Qiao, L.; Shi, F.; Liu, W.; Li, Y.; Dang, Y.P.; Gu, W.J.; Wang, X.G.; Liu, W. miR-98 suppresses melanoma metastasis through a negative feedback loop with its target gene IL-6. Exp. Mol. Med. 2014, 46, e116. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, W.; Chen, Z.; Wen, J.; Yu, H.; Wang, F.; Wu, L.; He, X. The effects of microRNA-98 inhibits cell proliferation and invasion by targeting STAT3 in nasopharyngeal carcinoma. Biomed. Pharmacother. 2017, 93, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, H.; Thomsen, A.R.; Haldrup, C.; Dyrskjot, L.; Hoyer, S.; Borre, M.; Mouritzen, P.; Orntoft, T.F.; Sorensen, K.D. Novel diagnostic and prognostic classifiers for prostate cancer identified by genome-wide microRNA profiling. Oncotarget 2016, 7, 30760–30771. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Li, J.; Ding, Q.; Cheng, G.; Zhou, H.; Tao, L.; Cai, H.; Li, P.; Cao, Q.; Ju, X.; et al. miR-152 controls migration and invasive potential by targeting TGFalpha in prostate cancer cell lines. Prostate 2013, 73, 1082–1089. [Google Scholar] [CrossRef] [PubMed]
- Theodore, S.C.; Davis, M.; Zhao, F.; Wang, H.; Chen, D.; Rhim, J.; Dean-Colomb, W.; Turner, T.; Ji, W.; Zeng, G.; et al. MicroRNA profiling of novel African American and Caucasian Prostate Cancer cell lines reveals a reciprocal regulatory relationship of miR-152 and DNA methyltranferase 1. Oncotarget 2014, 5, 3512–3525. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.W.; Zeng, J.; He, R.Q.; Rong, M.H.; Luo, D.Z.; Chen, G. Effects of miR-152 on cell growth inhibition, motility suppression and apoptosis induction in hepatocellular carcinoma cells. Asian Pac. J. Cancer Prev. 2014, 15, 4969–4976. [Google Scholar] [CrossRef]
- Wang, F.; Ying, H.; He, B.; Pan, Y.; Sun, H.; Wang, S. Circulating miR-148/152 family as potential biomarkers in hepatocellular carcinoma. Tumour Biol. 2016, 37, 4945–4953. [Google Scholar] [CrossRef]
- Zhang, P.; Sun, H.; Yang, B.; Luo, W.; Liu, Z.; Wang, J.; Zuo, Y. miR-152 regulated glioma cell proliferation and apoptosis via Runx2 mediated by DNMT1. Biomed. Pharmacother. 2017, 92, 690–695. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Liu, X.C.; Du, J.; Zhang, Y.J. MiR-152 regulates metastases of non-small cell lung cancer cells by targeting neuropilin-1. Int. J. Clin. Exp. Pathol. 2015, 8, 14235–14240. [Google Scholar] [PubMed]
- Dou, H.; Wang, Y.; Su, G.; Zhao, S. Decreased plasma let-7c and miR-152 as noninvasive biomarker for non-small-cell lung cancer. Int. J. Clin. Exp. Med. 2015, 8, 9291–9298. [Google Scholar] [PubMed]
- Chen, H.; Liu, H.; Zou, H.; Chen, R.; Dou, Y.; Sheng, S.; Dai, S.; Ai, J.; Melson, J.; Kittles, R.A.; et al. Evaluation of Plasma miR-21 and miR-152 as Diagnostic Biomarkers for Common Types of Human Cancers. J. Cancer 2016, 7, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Jiang, Y.; Yin, Y.; Li, Q.; He, J.; Jing, Y.; Qi, Y.T.; Xu, Q.; Li, W.; Lu, B.; et al. A regulatory circuit of miR-148a/152 and DNMT1 in modulating cell transformation and tumor angiogenesis through IGF-IR and IRS1. J. Mol. Cell Biol. 2013, 5, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Maimaitiming, A.; Wusiman, A.; Aimudula, A.; Tudahong, T.; Aisimutula, D. Downregulation of MicroRNA-152 and Inhibition of Cell Proliferation, Migration, and Invasion in Breast Cancer. Oncol. Res. 2017. [CrossRef] [PubMed]
- Li, B.; Xie, Z.; Li, B. miR-152 functions as a tumor suppressor in colorectal cancer by targeting PIK3R3. Tumour Biol. 2016, 37, 10075–10084. [Google Scholar] [CrossRef]
- Chen, Y.; Song, Y.; Wang, Z.; Yue, Z.; Xu, H.; Xing, C.; Liu, Z. Altered expression of MiR-148a and MiR-152 in gastrointestinal cancers and its clinical significance. J. Gastrointest. Surg. 2010, 14, 1170–1179. [Google Scholar] [CrossRef]
- Wang, Y.; Yuan, W.; Ma, X.; Ma, J. Expression of microRNA-152 in colorectal cancer and its relationship with prognosis. Zhonghua Zhong Liu Za Zhi 2016, 38, 763–766. [Google Scholar]
- Jiang, X.; Du, L.; Wang, L.; Li, J.; Liu, Y.; Zheng, G.; Qu, A.; Zhang, X.; Pan, H.; Yang, Y.; et al. Serum microRNA expression signatures identified from genome-wide microRNA profiling serve as novel noninvasive biomarkers for diagnosis and recurrence of bladder cancer. Int. J. Cancer 2015, 136, 854–862. [Google Scholar] [CrossRef]
- Dudziec, E.; Miah, S.; Choudhry, H.M.; Owen, H.C.; Blizard, S.; Glover, M.; Hamdy, F.C.; Catto, J.W. Hypermethylation of CpG islands and shores around specific microRNAs and mirtrons is associated with the phenotype and presence of bladder cancer. Clin. Cancer Res. 2011, 17, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhao, F.; Wang, Z.N.; Song, Y.X.; Chang, H.; Chiang, Y.; Xu, H.M. Altered expression of miR-152 and miR-148a in ovarian cancer is related to cell proliferation. Oncol. Rep. 2012, 27, 447–454. [Google Scholar] [PubMed]
- Zhai, R.; Kan, X.; Wang, B.; Du, H.; Long, Y.; Wu, H.; Tao, K.; Wang, G.; Bao, L.; Li, F.; et al. miR-152 suppresses gastric cancer cell proliferation and motility by targeting CD151. Tumour Biol. 2014, 35, 11367–11373. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Huang, C.; Li, S.; Yang, C.; Xi, Y.; Wang, L.; Zhang, F.; Fu, Y.; Li, D. Hsa-miR-326 targets CCND1 and inhibits non-small cell lung cancer development. Oncotarget 2016, 7, 8341–8359. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Hui, H.; Wang, L.J.; Wang, H.; Liu, Q.F.; Han, S.X. MicroRNA-326 functions as a tumor suppressor in colorectal cancer by targeting the nin one binding protein. Oncol. Rep. 2015, 33, 2309–2318. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Kong, F.; Deng, X.; Yu, Y.; Hou, C.; Liang, T.; Zhu, L. MicroRNA-326 suppresses the proliferation, migration and invasion of cervical cancer cells by targeting ELK1. Oncol. Lett. 2017, 13, 2949–2956. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Cao, X.; Chen, F. LncRNA-HOTAIR Activates Tumor Cell Proliferation and Migration by Suppressing MiR-326 in Cervical Cancer. Oncol. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gao, Y.; Xu, Y.; Ma, H.; Yang, M. Down-regulation of miR-326 is associated with poor prognosis and promotes growth and metastasis by targeting FSCN1 in gastric cancer. Growth Factors 2015, 33, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Wang, J.; Wang, P.Q. MiR-326 is a diagnostic biomarker and regulates cell survival and apoptosis by targeting Bcl-2 in osteosarcoma. Biomed. Pharmacother. 2016, 84, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Lin, S.; Hu, D.; Feng, Y.; Tan, Y.; Peng, Y. Interactions of miR-323/miR-326/miR-329 and miR-130a/miR-155/miR-210 as prognostic indicators for clinical outcome of glioblastoma patients. J. Transl. Med. 2013, 11, 10. [Google Scholar] [CrossRef]
- Wang, S.; Lu, S.; Geng, S.; Ma, S.; Liang, Z.; Jiao, B. Expression and clinical significance of microRNA-326 in human glioma miR-326 expression in glioma. Med. Oncol. 2013, 30, 373. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Li, Y.; Han, C.; Wang, X.; She, L.; Zhang, H. miRNA microarray reveals specific expression in the peripheral blood of glioblastoma patients. Int. J. Oncol. 2014, 45, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Goto, A.; Dobashi, Y.; Tsubochi, H.; Maeda, D.; Ooi, A. MicroRNAs associated with increased AKT gene number in human lung carcinoma. Hum. Pathol. 2016, 56, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wu, L.; Wang, A.; Tang, W.; Zhao, Y.; Zhao, H.; Teschendorff, A.E. dbDEMC 2.0: Updated database of differentially expressed miRNAs in human cancers. Nucleic Acids Res. 2017, 45, D812–D818. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Zhang, X.; Shi, J. MiR-98 inhibits cell proliferation and invasion of non-small cell carcinoma lung cancer by targeting PAK1. Int. J. Clin. Exp. Med. 2015, 8, 20135–20145. [Google Scholar] [PubMed]
- Wang, R.; Chen, X.; Xu, T.; Xia, R.; Han, L.; Chen, W.; De, W.; Shu, Y. MiR-326 regulates cell proliferation and migration in lung cancer by targeting phox2a and is regulated by HOTAIR. Am. J. Cancer Res. 2016, 6, 173–186. [Google Scholar] [PubMed]
- Hajjari, M.; Salavaty, A. HOTAIR: An oncogenic long non-coding RNA in different cancers. Cancer Biol. Med. 2015, 12, 1–9. [Google Scholar] [PubMed]
- Zhou, J.; Xu, T.; Yan, Y.; Qin, R.; Wang, H.; Zhang, X.; Huang, Y.; Wang, Y.; Lu, Y.; Fu, D.; et al. MicroRNA-326 functions as a tumor suppressor in glioma by targeting the Nin one binding protein (NOB1). PLoS ONE 2013, 8, e68469. [Google Scholar] [CrossRef] [PubMed]
- Biomarkers, Biomarkers and surrogate endpoints: Preferred definitions and conceptual framework. Clin. Pharmacol. Ther. 2001, 69, 89–95. [CrossRef] [PubMed]
- Lan, H.; Lu, H.; Wang, X.; Jin, H. MicroRNAs as Potential Biomarkers in Cancer: Opportunities and Challenges. BioMed. Res. Int. 2015, 2015, 125094. [Google Scholar] [CrossRef]
- Croce, C.M. Causes and consequences of microRNA dysregulation in cancer. Nat. Rev. Genet. 2009, 10, 704. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Croce, C.M. MicroRNA dysregulation in cancer: Diagnostics, monitoring and therapeutics. A comprehensive review. EMBO Mol. Med. 2012, 4, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Palanichamy, J.K.; Rao, D.S. miRNA dysregulation in cancer: Towards a mechanistic understanding. Front. Genet. 2014, 5, 54. [Google Scholar] [CrossRef] [PubMed]
- Park, N.J.; Zhou, H.; Elashoff, D.; Henson, B.S.; Kastratovic, D.A.; Abemayor, E.; Wong, D.T. Salivary microRNA: Discovery, characterization, and clinical utility for oral cancer detection. Clin. Cancer Res. 2009, 15, 5473–5477. [Google Scholar] [CrossRef] [PubMed]
- Hanke, M.; Hoefig, K.; Merz, H.; Feller, A.C.; Kausch, I.; Jocham, D.; Warnecke, J.M.; Sczakiel, G. A robust methodology to study urine microRNA as tumor marker: MicroRNA-126 and microRNA-182 are related to urinary bladder cancer. Urol. Oncol. 2010, 28, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Marzi, M.J.; Montani, F.; Carletti, R.M.; Dezi, F.; Dama, E.; Bonizzi, G.; Sandri, M.T.; Rampinelli, C.; Bellomi, M.; Maisonneuve, P.; et al. Optimization and Standardization of Circulating MicroRNA Detection for Clinical Application: The miR-Test Case. Clin. Chem. 2016, 62, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Filella, X.; Foj, L. miRNAs as novel biomarkers in the management of prostate cancer. Clin. Chem. Lab. Med. 2017, 55, 715–736. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Du, L.; Duan, W.; Wang, R.; Yan, K.; Wang, L.; Li, J.; Zheng, G.; Zhang, X.; Yang, Y.; et al. Serum microRNA expression signatures as novel noninvasive biomarkers for prediction and prognosis of muscle-invasive bladder cancer. Oncotarget 2016, 7, 36733–36742. [Google Scholar] [CrossRef] [PubMed]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or Tumor Suppressor? The Duplicity of MicroRNAs in Cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef]
- Daniel, R.; Wu, Q.; Williams, V.; Clark, G.; Guruli, G.; Zehner, Z. A Panel of MicroRNAs as Diagnostic Biomarkers for the Identification of Prostate Cancer. Int. J. Mol. Sci. 2017, 18, 1281. [Google Scholar] [CrossRef] [PubMed]
- Selth, L.A.; Townley, S.L.; Bert, A.G.; Stricker, P.D.; Sutherland, P.D.; Horvath, L.G.; Goodall, G.J.; Butler, L.M.; Tilley, W.D. Circulating microRNAs predict biochemical recurrence in prostate cancer patients. Br. J. Cancer 2013, 109, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Swanson, K.R.; True, L.D.; Lin, D.W.; Buhler, K.R.; Vessella, R.; Murray, J.D. A Quantitative Model for the Dynamics of Serum Prostate-Specific Antigen as a Marker for Cancerous Growth: An Explanation for a Medical Anomaly. Am. J. Pathol. 2001, 158, 2195–2199. [Google Scholar] [CrossRef]
- Kulis, M.; Esteller, M. DNA methylation and cancer. Adv. Genet. 2010, 70, 27–56. [Google Scholar] [PubMed]
- Klutstein, M.; Nejman, D.; Greenfield, R.; Cedar, H. DNA Methylation in Cancer and Aging. Cancer Res. 2016, 76, 3446–3450. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, J.; Qin, F.; Dai, S. miR-152 as a tumor suppressor microRNA: Target recognition and regulation in cancer. Oncol. Lett. 2016, 11, 3911–3916. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Lu, Z.; Unruh, A.K.; Ivan, C.; Baggerly, K.A.; Calin, G.A.; Li, Z.; Bast, R.C.; Le, X.-F. Clinically Relevant microRNAs in Ovarian Cancer. Mol. Cancer Res. MCR 2015, 13, 393–401. [Google Scholar] [CrossRef]
- Ramalho-Carvalho, J.; Gonçalves, C.S.; Graça, I.; Bidarra, D.; Pereira-Silva, E.; Salta, S.; Godinho, M.I.; Gomez, A.; Esteller, M.; Costa, B.M.; et al. A multiplatform approach identifies miR-152-3p as a common epigenetically regulated onco-suppressor in prostate cancer targeting TMEM97. Clin. Epigenet. 2018, 10, 40. [Google Scholar] [CrossRef]
- Özeş, A.R.; Wang, Y.; Zong, X.; Fang, F.; Pilrose, J.; Nephew, K.P. Therapeutic targeting using tumor specific peptides inhibits long non-coding RNA HOTAIR activity in ovarian and breast cancer. Sci. Rep. 2017, 7, 894. [Google Scholar] [CrossRef]
- National Cancer Institute. NCI Dictionary of Cancer Terms; National Cancer Institute: Bethesda, MD, USA, 2017. [Google Scholar]



{kind=link}
{kind=link}
| miRNA | Prostate | HCC | ESCC | Glioma | Lung | Breast | CRC | Bladder | Ovarian | Cervical | Gastric | Melanoma | NPC | OSC |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| miR-98 | ↑P [26,29]ᵒ | ↓T [30,31] | ↓T [32] | ↓T [33] | ↓T [34] ↓S [35] | ↓T [36] | ↓T [37] | |||||||
| miR-152 | ↑P [26] ↓T [38]# ↓T [39,40] | ↓T [41,42] | ↓T [43] | ↓T [44] ↓P [45] ↑P [46] | ↓T [47,48] ↑P [46] | ↓T [49,50,51] ↑P [46] | ↑S [52]↓T [53] | ↓T [54] | ↓T [50,55] | |||||
| miR-326 | ↑P [26] | ↓T [56] | ↓T [57] | ↓T [58,59] | ↓T [60] | ↓T/S [61] | ||||||||
| miR-4289 | ↑P [26] |
| miRNA | Cancer | Sample Type | Number of Samples | Hazard Ratio (HR) (95% Confidence Interval CI) | High %Survival | Mets. vs. No-Mets Tissue | Reference |
|---|---|---|---|---|---|---|---|
| miR-98-5p | Lung | Tissue | 26 | - | ↑ | ↓ | [35] |
| Melanoma | Tissue | 15 | - | ↑ | ↓ | [36] | |
| HCC | Tissue | 144 | - | ↑ | ↓ | [30] | |
| ESCC | Tissue | 40 | - | - | ↓ | [32] | |
| miR-152-3p | HCC | Serum | 76 | 0.39 (0.19–0.82) ᵒ | ↑ | - | [42] |
| Prostate | Tissue | 13 | - | ↑* | ↓ | [40] | |
| miR-326 | Prostate | Tissue | 126 | 1.1 (1.01–1.2) ᵒ,ᵞ | ↓ᵞ | - | [38] |
| Prostate | Tissue | 110 | 1.1 (1.01–1.2) ᵒ,ᵞ | ↓ᵞ | - | [38] | |
| CRC | Tissue | 114 | 0.58 (0.3–0.8) | ↑ | ↓ | [61] | |
| GBM | Tissue | 458 | 0.7 (0.5–0.9) | ↑ | - | [62] | |
| Glioma | Tissue | 108 | 0.15 (0.05–0.8) ᵒ | ↑ | - | [63] | |
| OSC | Serum | 60 | 0.25 (0.1–0.9) ᵒ | ↑ | ↓ | [61] | |
| Gastric | Tissue | 136 | 0.7 (0.4–0.9) ᵒ | ↑ | ↓ | [60] | |
| Lung | Tissue | 39 | - | ↑ | - | [56] |
| miRNA | Cancer | Target Gene/Regulator | Method of Validation | Binding Target Effect | Pathway or Molecular Function | Reference |
|---|---|---|---|---|---|---|
| miR-98 | Lung | Integrin Subunit Beta 3 (ITGB3) | Cancer cell line transfection and mouse injection model | Cancer cell proliferation suppression and tumor growth reduction in vivo | Cell adhesion and cell-surface mediated signalling | [34] |
| Serine/threonine-protein kinase (PAK1) | Cancer cell line transfection and mouse injection model | Cancer cell proliferation, colony formation, migration and invasion inhibition | Cytoskeleton reorganization and nuclear signalling | [67] | ||
| Melanoma | Interleukin 6 (IL-6) | Mouse injection model | Tumor metastasis and growth inhibition in vivo | Stat and NF-κB signalling | [36] | |
| HCC | Collagen Triple Helix Repeat Containing 1 (CTHRC1) | Western blot | Protein expression inhibition after mimics transfection | Tissue remodelling, vascularity and bone formation | [31] | |
| Brain | Raf-1 kinase inhibitor protein (RKIP) | Cancer cell line transfection | miR-98 positive regulator: tumor repressor | Inhibition of the Raf-1-MEK1/2, ERK1/2 and NF-kappaB signalling pathways | [33] | |
| NPC | Signal transducer and activator of transcription 3 (STAT3) | Western Blot | Protein expression inhibition after mimics transfection | Transcription factor | [37] | |
| miR-152 | Gastric | CD151 | Cancer cell line transfection | Cancer cell proliferation, migration and invasion suppression | Tetraspasin member (cell development, growth and motility regulation) | [55] |
| Ovarian | SERPINE1 | In silico transcriptome analysis | Tumorigenesis and metastasis suppression | tPA/uPA fibrinolysis inhibitor | [68] | |
| Breast | Insulin-like growth factor 1 (IGF-IR) | Cancer cell line transfection | Cancer cell proliferation, colony formation and tumor angiogenesis inhibition | PI3K/AKT and MAPK/ERK cascades | [47] | |
| Insulin receptor substrate 1 (IRS1) | Cancer cell line transfection | Cancer cell proliferation, colony formation, and tumor angiogenesis inhibition | PI3K/AKT and MAPK/ERK cascades | [47] | ||
| Rho-Associated Coiled-Coil Containing Protein Kinase 1 (ROCK1) | Cancer cell line transfection | Cancer cell proliferation, migration and invasion suppression | GTPase RhoA multiple signalling cascade | [48] | ||
| Lung | Neuropilin-1 mediated receptor | Cancer cell line transfection | Cancer cell migration and invasion suppression | VEGF-A, VEGF-165 | [54] | |
| miR-326 | Gastric | Fascin (FSCN1) | Cancer cell line transfection | Cancer cell growth and metastasis suppression | Formation of actin-based cellular protrusions | [60] |
| Lung | Cyclin D1 (CCND1) | Cancer cell line transfection | Cancer cell proliferation, migration, invasion and colony formation inhibition and promotion of apoptosis | CDK kinases regulator coordination of each mitotic event | [56] | |
| Paired-like homeobox 2a (phox2a) | Cancer cell line transfection | Cancer cell proliferation, migration and invasion suppression | Development of the autonomic nervous system | [69] | ||
| LncRNA-HOTAIR | Cancer cell line transfection | miR-326 negative regulator: promotion of cancer cells proliferation and migration | LSD1/PRC2 epigenetic regulator | [69] | ||
| Glioma | Nin one binding protein (NOB1) | Cancer cell line transfection | Cancer cell proliferation and colony formation inhibition | Pre-rRNA processing and MAPK signalling | [70] | |
| OSC | B-cell lymphoma 2 (Bcl-2) | Cancer cell line transfection | Cancer cell apoptosis reduction | Caspase activity regulator | [61] | |
| Cervical | LncRNA-HOTAIR | Cancer cell line transfection | miR-326 negative regulator: promotion of cancer cells proliferation and migration | LSD1/PRC2 epigenetic regulator | [59] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moya, L.; Meijer, J.; Schubert, S.; Matin, F.; Batra, J. Assessment of miR-98-5p, miR-152-3p, miR-326 and miR-4289 Expression as Biomarker for Prostate Cancer Diagnosis. Int. J. Mol. Sci. 2019, 20, 1154. https://doi.org/10.3390/ijms20051154
Moya L, Meijer J, Schubert S, Matin F, Batra J. Assessment of miR-98-5p, miR-152-3p, miR-326 and miR-4289 Expression as Biomarker for Prostate Cancer Diagnosis. International Journal of Molecular Sciences. 2019; 20(5):1154. https://doi.org/10.3390/ijms20051154
Chicago/Turabian StyleMoya, Leire, Jonelle Meijer, Sarah Schubert, Farhana Matin, and Jyotsna Batra. 2019. "Assessment of miR-98-5p, miR-152-3p, miR-326 and miR-4289 Expression as Biomarker for Prostate Cancer Diagnosis" International Journal of Molecular Sciences 20, no. 5: 1154. https://doi.org/10.3390/ijms20051154