Epigenetics: A Potential Mechanism Involved in the Pathogenesis of Various Adverse Consequences of Obstructive Sleep Apnea


Abstract
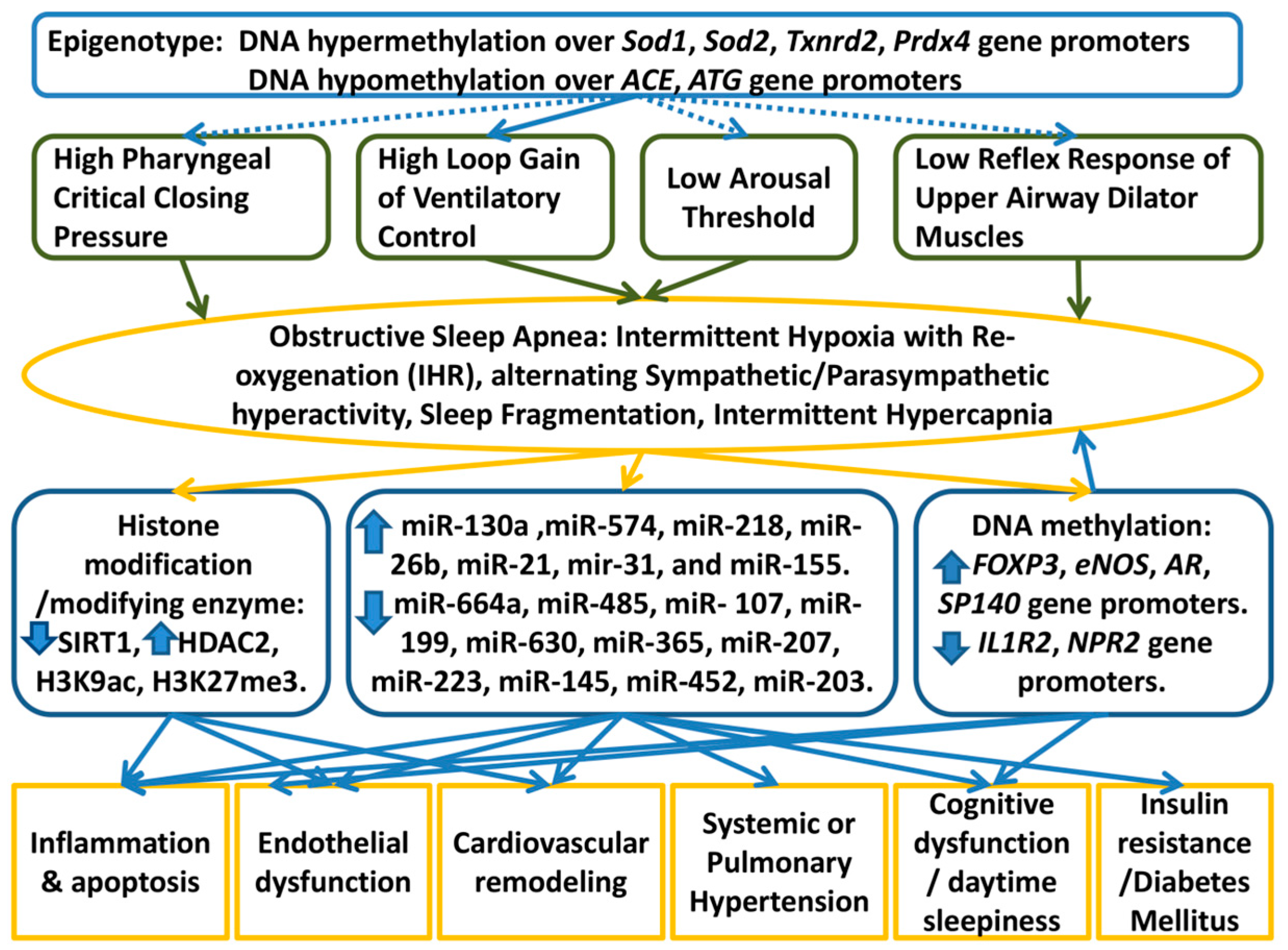
:1. Introduction
1.1. Epigenetic Mechanisms Underlying Sustained Hypoxia
1.2. Epigenetic Mechanisms Underlying IHR Injury in OSA
2. ROS-HIF-1-Endothelin and ROS-TLR-NF-κB Signaling Pathways in OSA
2.1. IHR Induces Systemic Inflammation and Sympathetic Hyperactivity through Enhancing ROS Production
2.2. ROS-HIF-1-Endothelin Signaling Pathway in OSA
2.3. ROS-TLR-NF-κB Signaling Pathway in OSA
3. Histone Modifications
3.1. Histone Modifications in Sustained Hypoxia
3.2. Abnormal Histone Modifying Enzyme Expressions in OSA Patients or in Response to IHR Experiments In Vitro
3.3. Histone Modification Patterns in Animals Exposed to CIH
4. Non-Coding RNAs
4.1. Differentially Expressed miRNAs in OSA Patients
4.2. Non-Coding RNA Changes in Animals Exposed to CIH or in Response to IHR In Vitro
5. DNA Methylation
5.1. Aberrant DNA Methylation Patterns in OSA Patients
5.2. Aberrant DNA Methylation Patterns in Animals Exposed to CIH
6. Crosstalk between microRNA and DNA Methylation/Histone Modification under Sustained Hypoxia Stimuli
7. Limitations and Perspectives of the OSA Cohort Studies, in vitro IHR Experiments, and CIH Animal Models
7.1. Limitations of Past Epigenetic Studies on OSA
7.2. Perspectives of Future Epigenetic-Related Investigations in OSA
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| OSA | obstructive sleep apnea |
| IHR | intermittent hypoxia with re-oxygenation |
| CPAP | continuous positive airway pressure |
| ROS | reactive oxygen species |
| HIF | hypoxia-inducible-factor |
| NF-κB | nuclear factor kappa B |
| AHI | apnea hypopnea index |
| miR | microRNA |
| lnc | long non-coding |
| NOX | NADPH oxidase |
| Sod | superoxide dismutase |
| ET-1 | endothelin |
| IL | interleukin |
| TNF | tumor necrosis factor |
| H | histone |
| ac | acetylation |
| me | methylation |
| HAT | histone acetyltransferase |
| HDAC | histone deacetylase |
| KDM | lysine demethylase |
| SIRT1 | sirtuin 1 |
| eNOS | endothelial nitric oxide synthase |
| CIH | chronic intermittent hypoxia |
| CpG | cytosine-phosphate-guanine |
| FOXP3 | forkhead box P3 |
| AR | androgen rceptor |
| NPR | natriuretic peptide receptor |
| SP140 | speckled protein140 |
| AOE | anti-oxidant enzyme |
| Txnrd | thioredoxin reductase |
| Prdx | peroxiredoxin |
| ACE | angiotensin converting enzyme |
| ATG | angiotensinogen |
| siRNA | small interfering RNA |
References
- Osman, A.M.; Carter, S.G.; Carberry, J.C.; Eckert, D.J. Obstructive sleep apnea: Current perspectives. Nat. Sci. Sleep 2018, 10, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Khalyfa, A.; Qiao, Z.; Gileles-Hillel, A.; Khalyfa, A.A.; Akbarpour, M.; Popko, B.; Gozal, D. Activation of the Integrated Stress Response and Metabolic Dysfunction in a Murine Model of Sleep Apnea. Am. J. Respir. Cell Mol. Biol. 2017, 57, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Gagnadoux, F.; Le Vaillant, M.; Paris, A.; Pigeanne, T.; Leclair-Visonneau, L.; Bizieux-Thaminy, A.; Alizon, C.; Humeau, M.P.; Nguyen, X.L.; Rouault, B.; et al. Relationship Between OSA Clinical Phenotypes and CPAP Treatment Outcomes. Chest 2016, 149, 288–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Zhou, Z.; McEvoy, R.D.; Anderson, C.S.; Rodgers, A.; Perkovic, V.; Neal, B. Association of Positive Airway Pressure with Cardiovascular Events and Death in Adults with Sleep Apnea: A Systematic Review and Meta-analysis. JAMA 2017, 318, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Cao, Q.; Guo, Z.; Dai, Q. Continuous Positive Airway Pressure in Patients with Obstructive Sleep Apnea and Resistant Hypertension: A Meta-Analysis of Randomized Controlled Trials. J. Clin. Hypertens. 2016, 18, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Iftikhar, I.H.; Khan, M.F.; Das, A.; Magalang, U.J. Meta-analysis: Continuous positive airway pressure improves insulin resistance in patients with sleep apnea without diabetes. Ann. Am. Thorac. Soc. 2013, 10, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Labarca, G.; Reyes, T.; Jorquera, J.; Dreyse, J.; Drake, L. CPAP in patients with obstructive sleep apnea and type 2 diabetes mellitus: Systematic review and meta-analysis. Clin. Respir. J. 2018, 12, 2361–2368. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Paulitsch, F.; Zhang, L. Continuous positive airway pressure for adults with obstructive sleep apnea and cardiovascular disease: A meta-analysis of randomized trials. Sleep Med. 2019, 54, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Camuzi, D.; de Amorim, I.S.S.; Ribeiro Pinto, L.F.; Oliveira Trivilin, L.; Mencalha, A.L.; Soares Lima, S.C. Regulation Is in the Air: The Relationship between Hypoxia and Epigenetics in Cancer. Cells 2019, 8, 300. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.H.; Ryan, J.J.; Marsboom, G.; Archer, S.L. Epigenetic mechanisms of pulmonary hypertension. Pulm. Circul. 2011, 1, 347–356. [Google Scholar] [CrossRef]
- Julian, C.G. Epigenomics and human adaptation to high altitude. J. Appl. Physiol. 2017, 123, 1362–1370. [Google Scholar] [CrossRef] [PubMed]
- Virzi, G.M.; Clementi, A.; Brocca, A.; de Cal, M.; Ronco, C. Epigenetics: A potential key mechanism involved in the pathogenesis of cardiorenal syndromes. J. Nephrol. 2018, 31, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Michiels, C.; Tellier, C.; Feron, O. Cycling hypoxia: A key feature of the tumor microenvironment. Biochim. Biophys. Acta 2016, 1866, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Tao, L.; Nie, P.; Lu, X.; Xu, X.; Chen, J.; Zhu, M. Association between 5-HT2A receptor polymorphisms and risk of obstructive sleep apnea and hypopnea syndrome: A systematic review and meta-analysis. Gene 2013, 530, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Varvarigou, V.; Dahabreh, I.J.; Malhotra, A.; Kales, S.N. A review of genetic association studies of obstructive sleep apnea: Field synopsis and meta-analysis. Sleep 2011, 34, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Redline, S.; Tishler, P.V.; Tosteson, T.D.; Williamson, J.; Kump, K.; Browner, I.; Ferrette, V.; Krejci, P. The familial aggregation of obstructive sleep apnea. Am. J. Respir. Crit. Care Med. 1995, 151, 682–687. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Cao, C.; Wu, Y.; Zhang, C.; Zhu, C.; Ying, S.; Chen, Z.; Shen, H.; Li, W. TNF-alpha-308G/A polymorphism contributes to obstructive sleep apnea syndrome risk: Evidence based on 10 case-control studies. PLoS ONE 2014, 9, e106183. [Google Scholar] [CrossRef]
- Lira, A.B.; de Sousa Rodrigues, C.F. Evaluation of oxidative stress markers in obstructive sleep apnea syndrome and additional antioxidant therapy: A review article. Sleep Breath. 2016, 20, 1155–1160. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, P.; Peng, Y.; Ouyang, R. Role of Oxidative Stress in the Neurocognitive Dysfunction of Obstructive Sleep Apnea Syndrome. Oxid. Med. Cell. Longev. 2016, 2016, 9626831. [Google Scholar] [CrossRef]
- Choudhry, H.; Harris, A.L. Advances in Hypoxia-Inducible Factor Biology. Cell Metab. 2018, 27, 281–298. [Google Scholar] [CrossRef]
- Nanduri, J.; Peng, Y.J.; Yuan, G.; Kumar, G.K.; Prabhakar, N.R. Hypoxia-inducible factors and hypertension: Lessons from sleep apnea syndrome. J. Mol. Med. 2015, 93, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L.; Prabhakar, N.R. Neural regulation of hypoxia-inducible factors and redox state drives the pathogenesis of hypertension in a rodent model of sleep apnea. J. Appl. Physiol. 2015, 119, 1152–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gileles-Hillel, A.; Almendros, I.; Khalyfa, A.; Nigdelioglu, R.; Qiao, Z.; Hamanaka, R.B.; Mutlu, G.M.; Akbarpour, M.; Gozal, D. Prolonged Exposures to Intermittent Hypoxia Promote Visceral White Adipose Tissue Inflammation in a Murine Model of Severe Sleep Apnea: Effect of Normoxic Recovery. Sleep 2017, 40. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.L.; Gozal, D.; Wang, Y.; Bandla, H.P.; Bhattacharjee, R.; Kulkarni, R.; Kheirandish-Gozal, L. Alterations in circulating T-cell lymphocyte populations in children with obstructive sleep apnea. Sleep 2013, 36, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Schulz, R.; Mahmoudi, S.; Hattar, K.; Sibelius, U.; Olschewski, H.; Mayer, K.; Seeger, W.; Grimminger, F. Enhanced release of superoxide from polymorphonuclear neutrophils in obstructive sleep apnea. Impact of continuous positive airway pressure therapy. Am. J. Respir. Crit. Care Med. 2000, 162, 566–570. [Google Scholar] [CrossRef] [PubMed]
- Dyugovskaya, L.; Polyakov, A.; Lavie, P.; Lavie, L. Delayed neutrophil apoptosis in patients with sleep apnea. Am. J. Respir. Crit. Care Med. 2008, 177, 545–554. [Google Scholar] [CrossRef]
- Khalyfa, A.; Kheirandish-Gozal, L.; Khalyfa, A.A.; Philby, M.F.; Alonso-Alvarez, M.L.; Mohammadi, M.; Bhattacharjee, R.; Teran-Santos, J.; Huang, L.; Andrade, J.; et al. Circulating Plasma Extracellular Microvesicle MicroRNA Cargo and Endothelial Dysfunction in Children with Obstructive Sleep Apnea. Am. J. Respir. Crit. Care Med. 2016, 194, 1116–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.C.; Chen, K.D.; Su, M.C.; Chin, C.H.; Chen, C.J.; Liou, C.W.; Chen, T.W.; Chang, Y.C.; Huang, K.T.; Wang, C.C.; et al. Genome-wide gene expression array identifies novel genes related to disease severity and excessive daytime sleepiness in patients with obstructive sleep apnea. PLoS ONE 2017, 12, e0176575. [Google Scholar] [CrossRef]
- Schaefer, E.; Wu, W.; Mark, C.; Yang, A.; DiGiacomo, E.; Carlton-Smith, C.; Salloum, S.; Brisac, C.; Lin, W.; Corey, K.E.; et al. Intermittent hypoxia is a proinflammatory stimulus resulting in IL-6 expression and M1 macrophage polarization. Hepatol. Commun. 2017, 1, 326–337. [Google Scholar] [CrossRef]
- Chen, Y.C.; Su, M.C.; Chin, C.H.; Lin, I.C.; Hsu, P.Y.; Liou, C.W.; Huang, K.T.; Wang, T.Y.; Lin, Y.Y.; Zheng, Y.X.; et al. Formyl peptide receptor 1 up-regulation and formyl peptide receptor 2/3 down-regulation of blood immune cells along with defective lipoxin A4/resolvin D1 production in obstructive sleep apnea patients. PLoS ONE 2019, 14, e0216607. [Google Scholar] [CrossRef]
- Belaidi, E.; Morand, J.; Gras, E.; Pepin, J.L.; Godin-Ribuot, D. Targeting the ROS-HIF-1-endothelin axis as a therapeutic approach for the treatment of obstructive sleep apnea-related cardiovascular complications. Pharmacol. Ther. 2016, 168, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Janssen, C.; Pathak, A.; Grassi, G.; van de Borne, P. Endothelin contributes to the blood pressure rise triggered by hypoxia in severe obstructive sleep apnea. J. Hypertens. 2017, 35, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.J.; Wang, S.J.; Gao, X.; Wang, B.; Dong, Y.T.; Bai, Y.; Chen, Y.; Gong, J.N.; Huang, Y.Q.; An, D.D. Toll-Like Receptor 4 (TLR-4) Pathway Promotes Pulmonary Inflammation in Chronic Intermittent Hypoxia-Induced Obstructive Sleep Apnea. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 7152–7161. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Guo, R.; Dong, M.; Zheng, J.; Lin, H.; Lu, H. Contribution of TLR4 signaling in intermittent hypoxia-mediated atherosclerosis progression. J. Transl. Med. 2018, 16, 106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Su, X.; Zou, F.; Xu, T.; Pan, P.; Hu, C. Toll-like receptor-4 deficiency alleviates chronic intermittent hypoxia-induced renal injury, inflammation, and fibrosis. Sleep Breath. 2018. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Su, M.C.; Liou, C.W.; Liu, S.F.; Chen, C.J.; Lin, H.C.; Hsiao, C.C.; Wang, T.Y.; Wang, C.C.; Chin, C.H.; et al. Co-upregulation of Toll-like receptors 2 and 6 on peripheral blood cells in patients with obstructive sleep apnea. Sleep Breath. 2015, 19, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Amin, Z.; Amin, H.Z.; Amin, L.Z. Obstructive Sleep Apnea and Atherosclerosis. Acta Med. Indones. 2016, 48, 636–637. [Google Scholar]
- Collins, P.L.; Oltz, E.M. Histone methylation keeps the brakes on autophagy. Mol. Cell. Biol. 2013, 33, 3974–3975. [Google Scholar] [CrossRef]
- Watson, J.A.; Watson, C.J.; McCrohan, A.M.; Woodfine, K.; Tosetto, M.; McDaid, J.; Gallagher, E.; Betts, D.; Baugh, J.; O’Sullivan, J.; et al. Generation of an epigenetic signature by chronic hypoxia in prostate cells. Human Mol. Genet. 2009, 18, 3594–3604. [Google Scholar] [CrossRef]
- Benlhabib, H.; Mendelson, C.R. Epigenetic regulation of surfactant protein A gene (SP-A) expression in fetal lung reveals a critical role for Suv39h methyltransferases during development and hypoxia. Mol. Cell. Biol. 2011, 31, 1949–1958. [Google Scholar] [CrossRef]
- Bouquet, F.; Ousset, M.; Biard, D.; Fallone, F.; Dauvillier, S.; Frit, P.; Salles, B.; Muller, C. A DNA-dependent stress response involving DNA-PK occurs in hypoxic cells and contributes to cellular adaptation to hypoxia. J. Cell Sci. 2011, 124, 1943–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Chu, A.; Turker, M.S.; Glazer, P.M. Hypoxia-induced epigenetic regulation and silencing of the BRCA1 promoter. Mol. Cell. Biol. 2011, 31, 3339–3350. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, K.; Vlachojannis, G.J.; Koyanagi, M.; Boeckel, J.N.; Urbich, C.; Farcas, R.; Bonig, H.; Marquez, V.E.; Zeiher, A.M.; Dimmeler, S. Epigenetic regulation of endothelial lineage committed genes in pro-angiogenic hematopoietic and endothelial progenitor cells. Circul. Res. 2011, 109, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Yang, E.G.; Park, H. Hypoxia enhances the expression of prostate-specific antigen by modifying the quantity and catalytic activity of Jumonji C domain-containing histone demethylases. Carcinogenesis 2013, 34, 2706–2715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dmitriev, R.I.; Papkovsky, D.B. In vitro ischemia decreases histone H4K16 acetylation in neural cells. FEBS Lett. 2015, 589, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zhang, L.; Zhuang, S.; Qin, G.J.; Zhao, T.C. HDAC4 degradation mediates HDAC inhibition-induced protective effects against hypoxia/reoxygenation injury. J. Cell. Physiol. 2015, 230, 1321–1331. [Google Scholar] [CrossRef] [PubMed]
- Safronova, O.S.; Nakahama, K.; Morita, I. Acute hypoxia affects P-TEFb through HDAC3 and HEXIM1-dependent mechanism to promote gene-specific transcriptional repression. Nucleic Acids Res. 2014, 42, 8954–8969. [Google Scholar] [CrossRef]
- Hong, F.; Wan, L.; Liu, J.; Huang, K.; Xiao, Z.; Zhang, Y.; Shi, C. Histone methylation regulates Hif-1 signaling cascade in activation of hepatic stellate cells. FEBS Open Bio 2018, 8, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Nanduri, J.; Peng, Y.J.; Wang, N.; Khan, S.A.; Semenza, G.L.; Kumar, G.K.; Prabhakar, N.R. Epigenetic regulation of redox state mediates persistent cardiorespiratory abnormalities after long-term intermittent hypoxia. J. Physiol. 2017, 595, 63–77. [Google Scholar] [CrossRef]
- Roman, J.; Zhu, J.; Ritzenthaler, J.D.; Zelko, I.N. Epigenetic regulation of EC-SOD expression in aging lung fibroblasts: Role of histone acetylation. Free Radic. Biol. Med. 2017, 112, 212–223. [Google Scholar] [CrossRef]
- Chen, F.; Li, X.; Aquadro, E.; Haigh, S.; Zhou, J.; Stepp, D.W.; Weintraub, N.L.; Barman, S.A.; Fulton, D.J.R. Inhibition of histone deacetylase reduces transcription of NADPH oxidases and ROS production and ameliorates pulmonary arterial hypertension. Free Radic. Biol. Med. 2016, 99, 167–168. [Google Scholar] [CrossRef] [PubMed]
- Manea, S.A.; Antonescu, M.L.; Fenyo, I.M.; Raicu, M.; Simionescu, M.; Manea, A. Epigenetic regulation of vascular NADPH oxidase expression and reactive oxygen species production by histone deacetylase-dependent mechanisms in experimental diabetes. Redox Biol. 2018, 16, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Dobrynin, G.; McAllister, T.E.; Leszczynska, K.B.; Ramachandran, S.; Krieg, A.J.; Kawamura, A.; Hammond, E.M. KDM4A regulates HIF-1 levels through H3K9me3. Sci. Rep. 2017, 7, 11094. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.T.; Wang, H.; Zhang, S.; Du, J.; Zhuang, S.; Zhao, T.C. Irisin Ameliorates Hypoxia/Reoxygenation-Induced Injury through Modulation of Histone Deacetylase 4. PLoS ONE 2016, 11, e0166182. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Wang, H.Y.; Liaw, S.F.; Chiu, C.H.; Lin, M.W. Effect of oral appliance on circulating leukocyte telomere length and SIRT1 in obstructive sleep apnea. Clin. Oral Investig. 2018. [Google Scholar] [CrossRef]
- Chen, W.J.; Liaw, S.F.; Lin, C.C.; Chiu, C.H.; Lin, M.W.; Chang, F.T. Effect of Nasal CPAP on SIRT1 and Endothelial Function in Obstructive Sleep Apnea Syndrome. Lung 2015, 193, 1037–1045. [Google Scholar] [CrossRef]
- Chen, J.H.; Yang, R.; Wang, Y.P.; Zhang, W. Expression data analysis to identify key target genes in visceral fat tissue associated with obstructive sleep apnea. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4293–4299. [Google Scholar]
- Cortese, R.; Gileles-Hillel, A.; Khalyfa, A.; Almendros, I.; Akbarpour, M.; Khalyfa, A.A.; Qiao, Z.; Garcia, T.; Andrade, J.; Gozal, D. Aorta macrophage inflammatory and epigenetic changes in a murine model of obstructive sleep apnea: Potential role of CD36. Sci. Rep. 2017, 7, 43648. [Google Scholar] [CrossRef] [Green Version]
- Joosten, S.C.; Smits, K.M.; Aarts, M.J.; Melotte, V.; Koch, A.; Tjan-Heijnen, V.C.; van Engeland, M. Epigenetics in renal cell cancer: Mechanisms and clinical applications. Nat. Rev. Urol. 2018, 15, 430–451. [Google Scholar] [CrossRef]
- Sanchez-de-la-Torre, M.; Khalyfa, A.; Sanchez-de-la-Torre, A.; Martinez-Alonso, M.; Martinez-Garcia, M.A.; Barcelo, A.; Lloberes, P.; Campos-Rodriguez, F.; Capote, F.; Diaz-de-Atauri, M.J.; et al. Precision Medicine in Patients with Resistant Hypertension and Obstructive Sleep Apnea: Blood Pressure Response to Continuous Positive Airway Pressure Treatment. J. Am. Coll. Cardiol. 2015, 66, 1023–1032. [Google Scholar] [CrossRef]
- Li, K.; Chen, Z.; Qin, Y.; Wei, Y. MiR-664a-3p expression in patients with obstructive sleep apnea: A potential marker of atherosclerosis. Medicine 2018, 97, e9813. [Google Scholar] [CrossRef] [PubMed]
- An, Z.; Wang, D.; Yang, G.; Zhang, W.Q.; Ren, J.; Fu, J.L. Role of microRNA-130a in the pathogeneses of obstructive sleep apnea hypopnea syndrome-associated pulmonary hypertension by targeting the GAX gene. Medicine 2017, 96, e6746. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Wei, P.; Qin, Y.; Wei, Y. MicroRNA expression profiling and bioinformatics analysis of dysregulated microRNAs in obstructive sleep apnea patients. Medicine 2017, 96, e7917. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.X.; Chen, G.P.; Lin, P.L.; Huang, J.C.; Lin, X.; Qi, J.C.; Lin, Q.C. Detection and analysis of apoptosis- and autophagy-related miRNAs of mouse vascular endothelial cells in chronic intermittent hypoxia model. Life Sci. 2018, 193, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.X.; Chen, Q.; Chen, G.P.; Huang, J.C.; Huang, J.F.; He, X.R.; Lin, T.; Lin, Q.C. Inhibition of microRNA-218 reduces HIF-1alpha by targeting on Robo1 in mice aortic endothelial cells under intermittent hypoxia. Oncotarget 2017, 8, 104359–104366. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Han, Z.; Huang, S.; Bai, R.; Ge, X.; Chen, F.; Lei, P. Intermittent hypoxia caused cognitive dysfunction relate to miRNAs dysregulation in hippocampus. Behav. Brain Res. 2017, 335, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, X.; Li, B.; Zhang, Q.; Wang, J.; Zhang, W.; Luo, W.; Chen, J. Cold Inducible RNA Binding Protein Is Involved in Chronic Hypoxia Induced Neuron Apoptosis by Down-Regulating HIF-1alpha Expression and Regulated by microRNA-23a. Int. J. Biol. Sci. 2017, 13, 518–531. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.; Jiang, L.; Fu, C.; Wu, X.; Liu, Z.; Song, J.; Lu, H.; Wu, X.; Li, S. 2-Methoxyestradiol attenuates chronic-intermittent-hypoxia-induced pulmonary hypertension through regulating microRNA-223. J. Cell. Physiol. 2019, 234, 6324–6335. [Google Scholar] [CrossRef]
- Zhang, K.; Ma, Z.; Wang, W.; Liu, R.; Zhang, Y.; Yuan, M.; Li, G. Beneficial effects of tolvaptan on atrial remodeling induced by chronic intermittent hypoxia in rats. Cardiovasc. Ther. 2018, 36, e12466. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, K.; Li, X.; Ma, Z.; Zhang, Y.; Yuan, M.; Suo, Y.; Liang, X.; Tse, G.; Goudis, C.A.; et al. Doxycycline attenuates chronic intermittent hypoxia-induced atrial fibrosis in rats. Cardiovasc. Ther. 2018, 36, e12321. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Liu, W.; Li, G.C.; Jin, M.; You, Z.X.; Liu, H.G.; Hu, Y. Atorvastatin Attenuates Myocardial Hypertrophy Induced by Chronic Intermittent Hypoxia In Vitro Partly through miR-31/PKCepsilon Pathway. Curr. Med. Sci. 2018, 38, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Chang, S.C.; Jin, J.; Gu, W.; Li, S. NLRP3 in fl ammasome mediates chronic intermittent hypoxia-induced renal injury implication of the microRNA-155/FOXO3a signaling pathway. J. Cell. Physiol. 2018, 233, 9404–9415. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, T.; Itaya-Hironaka, A.; Yamauchi, A.; Makino, M.; Sakuramoto-Tsuchida, S.; Shobatake, R.; Ota, H.; Takeda, M.; Ohbayashi, C.; Takasawa, S. Intermittent Hypoxia Up-Regulates CCL2, RETN, and TNFalpha mRNAs in Adipocytes via Down-regulation of miR-452. Int. J. Mol. Sci. 2019, 20, 1960. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, T.; Ota, H.; Itaya-Hironaka, A.; Shobatake, R.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Makino, M.; Kimura, H.; Takeda, M.; Ohbayashi, C.; et al. Up-regulation of selenoprotein P and HIP/PAP mRNAs in hepatocytes by intermittent hypoxia via down-regulation of miR-203. Biochem. Biophys. Rep. 2017, 11, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Lin, G.; Huang, J.; Chen, G.; Huang, X.; Lin, Q. Expression profile of long non-coding RNAs in rat models of OSA-induced cardiovascular disease: New insight into pathogenesis. Sleep Breath. 2018. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Liu, Y.; Sun, L.; Wang, D.; Wang, Y.; Zhao, S.; Dai, H.; Zhao, J.; Zhang, S.; Li, M.; et al. Chronic obstructive sleep apnea promotes aortic remodeling in canines through miR-145/Smad3 signaling pathway. Oncotarget 2017, 8, 37705–37716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maunakea, A.K.; Nagarajan, R.P.; Bilenky, M.; Ballinger, T.J.; D’Souza, C.; Fouse, S.D.; Johnson, B.E.; Hong, C.; Nielsen, C.; Zhao, Y.; et al. Conserved role of intragenic DNA methylation in regulating alternative promoters. Nature 2010, 466, 253–257. [Google Scholar] [CrossRef]
- Luo, C.; Hajkova, P.; Ecker, J.R. Dynamic DNA methylation: In the right place at the right time. Science 2018, 361, 1336–1340. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Bhattacharjee, R.; Khalyfa, A.; Kheirandish-Gozal, L.; Capdevila, O.S.; Wang, Y.; Gozal, D. DNA methylation in inflammatory genes among children with obstructive sleep apnea. Am. J. Respir. Crit. Care Med. 2012, 185, 330–338. [Google Scholar] [CrossRef]
- Kheirandish-Gozal, L.; Khalyfa, A.; Gozal, D.; Bhattacharjee, R.; Wang, Y. Endothelial dysfunction in children with obstructive sleep apnea is associated with epigenetic changes in the eNOS gene. Chest 2013, 143, 971–977. [Google Scholar] [CrossRef]
- Chen, Y.C.; Chen, T.W.; Su, M.C.; Chen, C.J.; Chen, K.D.; Liou, C.W.; Tang, P.; Wang, T.Y.; Chang, J.C.; Wang, C.C.; et al. Whole Genome DNA Methylation Analysis of Obstructive Sleep Apnea: IL1R2, NPR2, AR, SP140 Methylation and Clinical Phenotype. Sleep 2016, 39, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Nanduri, J.; Makarenko, V.; Reddy, V.D.; Yuan, G.; Pawar, A.; Wang, N.; Khan, S.A.; Zhang, X.; Kinsman, B.; Peng, Y.J.; et al. Epigenetic regulation of hypoxic sensing disrupts cardiorespiratory homeostasis. Proc. Natl. Acad. Sci. USA 2012, 109, 2515–2520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, A.; Gozal, D.; Cortese, R.; Wang, Y. Cardiovascular dysfunction in adult mice following postnatal intermittent hypoxia. Pediatr. Res. 2015, 77, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Cortese, R.; Almendros, I.; Wang, Y.; Gozal, D. Tumor circulating DNA profiling in xenografted mice exposed to intermittent hypoxia. Oncotarget 2015, 6, 556–569. [Google Scholar] [CrossRef]
- Perikleous, E.; Steiropoulos, P.; Tzouvelekis, A.; Nena, E.; Koffa, M.; Paraskakis, E. DNA Methylation in Pediatric Obstructive Sleep Apnea: An Overview of Preliminary Findings. Front. Pediatr. 2018, 6, 154. [Google Scholar] [CrossRef] [Green Version]
- Strmsek, Z.; Kunej, T. MicroRNA Silencing by DNA Methylation in Human Cancer: A Literature Analysis. Non-Coding RNA 2015, 1, 44–52. [Google Scholar] [CrossRef]
- Xiong, L.; Wang, F.; Huang, X.; Liu, Z.H.; Zhao, T.; Wu, L.Y.; Wu, K.; Ding, X.; Liu, S.; Wu, Y.; et al. DNA demethylation regulates the expression of miR-210 in neural progenitor cells subjected to hypoxia. FEBS J. 2012, 279, 4318–4326. [Google Scholar] [CrossRef]
- Doridot, L.; Houry, D.; Gaillard, H.; Chelbi, S.T.; Barbaux, S.; Vaiman, D. miR-34a expression, epigenetic regulation, and function in human placental diseases. Epigenetics 2014, 9, 142–151. [Google Scholar] [CrossRef]
- Gong, M.; Liang, T.; Jin, S.; Dai, X.; Zhou, Z.; Gao, M.; Huang, S.; Luo, J.; Zou, L.; Zou, X. Methylation-mediated silencing of miR-124 facilitates chondrogenesis by targeting NFATc1 under hypoxic conditions. Am. J. Transl. Res. 2017, 9, 4111–4124. [Google Scholar]
- Zinchuk, A.V.; Gentry, M.J.; Concato, J.; Yaggi, H.K. Phenotypes in obstructive sleep apnea: A definition, examples and evolution of approaches. Sleep Med. Rev. 2016. [Google Scholar] [CrossRef]
- Shanks, N.; Greek, R.; Greek, J. Are animal models predictive for humans? Philos. Ethics Hum. Med. 2009, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Schultz, M.D.; He, Y.; Whitaker, J.W.; Hariharan, M.; Mukamel, E.A.; Leung, D.; Rajagopal, N.; Nery, J.R.; Urich, M.A.; Chen, H.; et al. Human body epigenome maps reveal noncanonical DNA methylation variation. Nature 2015, 523, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Cobos, S.N.; Bennett, S.A.; Torrente, M.P. The impact of histone post-translational modifications in neurodegenerative diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2018. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Doss, C.G.P.; Lee, S.S. Therapeutic miRNA and siRNA: Moving from Bench to Clinic as Next Generation Medicine. Mol. Ther. Nucleic Acids 2017, 8, 132–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Histone Modification or Modifying Enzyme | Attribute | Up- or Down-Regulation in Response to IHR | Investigation Models | Main Role of the Histone Modification or Modifying Enzyme | Reference |
|---|---|---|---|---|---|
| SIRT1 | Class III HDAC | Down | Peripheral blood mononuclear cells of OSA patients | Repress endothelial nitric oxide synthase; negatively correlated with AHI and minimum SaO2 | [55,56] |
| HDAC2 | Class I HDAC | Up | Visceral fat of OSA patients | [57] | |
| H3K9Ac | Active histone mark | Increased enrichment over Foxo4, Lrtm2, Gabbr2, ABca1, IL6 | Aortic macrophages of rats, CIH for 20 weeks | Activate pro-inflammatory genes | [58] |
| H3K27me3 | Repressive histone mark | Increased enrichment over PPAR/RXR and LXR/RXR | Aortic macrophages of rats, CIH for 20 weeks | Repress anti-inflammatory genes | [58] |
| Up- or Down-Regulation in Response to IHR | Investigation Models | Main Role of the miR | TARGET GENE * | Reference | |
|---|---|---|---|---|---|
| miR-378a-3p | up | Plasma of OSA patients with resistant hypertension | Predict blood pressure decreases to CPAP treatment | [60] | |
| miR-100-5p | up | Plasma of OSA patients with resistant hypertension | Predict blood pressure decreases to CPAP treatment | [60] | |
| miR-486-5p | down | Plasma of OSA patients with resistant hypertension | Predict blood pressure decreases to CPAP treatment | [60] | |
| miR-664a | down | Serum of OSA patients | A marker of atherosclerosis; negatively correlated with AHI | [61] | |
| miR-130a | up | Blood of OSA patients; human umbilical vein endothelial cell | Potentiate pulmonary hypertension | GAX | [62] |
| miR-485-5p | down | Serum of OSA patients | [63] | ||
| miR-107 | down | Serum of OSA patients | [63] | ||
| miR-199-3p | down | Serum of OSA patients | [63] | ||
| miR-574-5p | up | Serum of OSA patients | [63] | ||
| miR-630 | down | Pediatric OSA patients; human microvascular endothelial cells | Attenuate endothelial dysfunction | Nrf2, AMP kinase, and tight junction pathways | [27] |
| miR-223 | down | Rats, CIH for 6 weeks | Attenuate pulmonary hypertension | [68] | |
| miR-21 | up | Rats, CIH for 30 days | Induce atrial remodeling and fibrosis | Spry1/ERK/MMP-9; PTEN/PI3K/AKT | [69,70] |
| miR-155 | up | Mice, CIH for >4 weeks; in vitro HK-2 cells, IHR | Promote kidney injury | FOXO3a | [72] |
| miR-31 | up | In vitro H9c2 cardiomyocyte, IHR | Promote cardiac hypertrophy | PKCε | [71] |
| miR-145 | down | Canines, CIH for 12 weeks | Attenuate aortic remodeling and sympathetic nerve sprouting | Smad3 | [76] |
| miR-365 | down | In vitro Hepatocyte/macrophage, IHR | Inhibit inflammation | IL-6 | [29] |
| miR-218 | up | In vitro mice aortic endothelium, IHR | Potentiate apoptosis | Robo1 | [65] |
| miR-26b | up | Rats, CIH for 4 weeks | Promote cognitive dysfunction | [66] | |
| miR-207 | down | Rats, CIH for 4 weeks | Attenuate cognitive dysfunction | [66] | |
| miR-452 | down | mouse 3T3-L1 and human SW872 adipocytes, IHR | Attenuate insulin resistance | RETN, TNF-α, and CCL2 | [73] |
| miR-203 | down | Human JHH5, JHH7, and HepG2 hepatocytes, IHR | Potentiate insulin resistance | SELENOP HIP/PAP | [74] |
| Genes | Hyper- or Hypo-Methylation in OSA or in Response to IHR | Investigation Model | Main Role of the Aberrant DNA Methylation | References |
|---|---|---|---|---|
| FOXP3 | Hypermethylated intron1 region (mean of 11 CpG sites) | Pediatric OSA patients with high hypersensitivity CRP | Positively correlated with AHI and hypersensitivity CRP | [79] |
| eNOS | Hypermethylated promoter region (-171 CpG site) | Pediatric OSA patients with endothelial dysfunction | Decreased eNOS gene expression | [80] |
| IL1R2 | Hypomethylated promoter region (-114 CpG site) | Adult OSA patients | Increased IL1R2 protein expression; negatively correlated with oxygen desaturation index | [81] |
| AR | Hypermethylated promoter region (-531 CpG site) | Adult OSA patients | Positively correlated with AHI | [81] |
| NPR2 | Hypomethylated promoter region (-608/-618 CpG sites) | Adult OSA patients with excessive daytime sleepiness | Increased NPR2 and CNP protein expressions; negatively correlated with Epworth Sleepiness Scale | [81] |
| SP140 | Hypermethylated promoter region (-194 CpG site) | Adult OSA patients with excessive daytime sleepiness | Decreased SP140 protein expression; positively correlated with Epworth Sleepiness Scale | [81] |
| Anti-oxidant enzymes (AOE genes: Sod1, Sod2, Txnrd2, Prdx4) | Hypermethylated promoter regions | Rats, CIH for 30 days; neonatal rats, IH from postnatal day 1 to day 10 | Inhibit AOE genes, increase ROS production, exaggerate chemoreflexes of the carotid body | [49,82] |
| Rab3a | Hypermethylated promoter regions | Mice engrafted with TC1 epithelial lung cancer cells, CIH for 2 weeks followed by tumor engraftment for 4 weeks | Increased tumor growth and invasion | [84] |
| Ace1 | Hypomethylated promoter regions | mesenteric endothelial cells of mice, CIH for the first 4 weeks of life | Increased Ace gene expression; diminished vasodilatory responses, increased ROS content | [83] |
| Atg | Hypomethylated enhancer regions | mesenteric endothelial cells of mice, CIH for the first 4 weeks of life | Increased angiotensinogen (Atg) protein expression; diminished vasodilatory responses, increased ROS content | [83] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-C.; Hsu, P.-Y.; Hsiao, C.-C.; Lin, M.-C. Epigenetics: A Potential Mechanism Involved in the Pathogenesis of Various Adverse Consequences of Obstructive Sleep Apnea. Int. J. Mol. Sci. 2019, 20, 2937. https://doi.org/10.3390/ijms20122937
Chen Y-C, Hsu P-Y, Hsiao C-C, Lin M-C. Epigenetics: A Potential Mechanism Involved in the Pathogenesis of Various Adverse Consequences of Obstructive Sleep Apnea. International Journal of Molecular Sciences. 2019; 20(12):2937. https://doi.org/10.3390/ijms20122937
Chicago/Turabian StyleChen, Yung-Che, Po-Yuan Hsu, Chang-Chun Hsiao, and Meng-Chih Lin. 2019. "Epigenetics: A Potential Mechanism Involved in the Pathogenesis of Various Adverse Consequences of Obstructive Sleep Apnea" International Journal of Molecular Sciences 20, no. 12: 2937. https://doi.org/10.3390/ijms20122937