Effects on the Profile of Circulating miRNAs after Single Bouts of Resistance Training with and without Blood Flow Restriction—A Three-Arm, Randomized Crossover Trial

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Sample
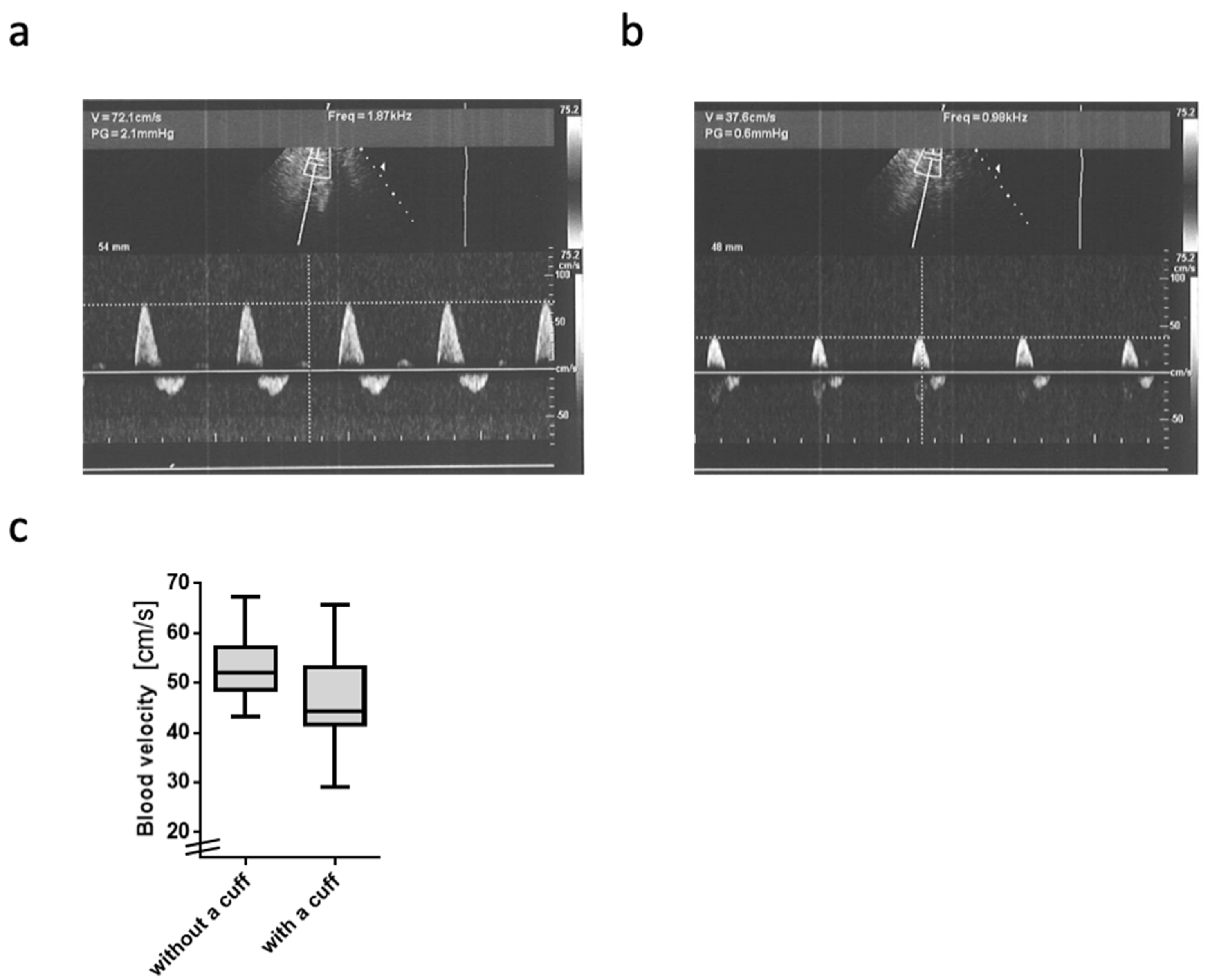
2.2. Blood Flow Velocity
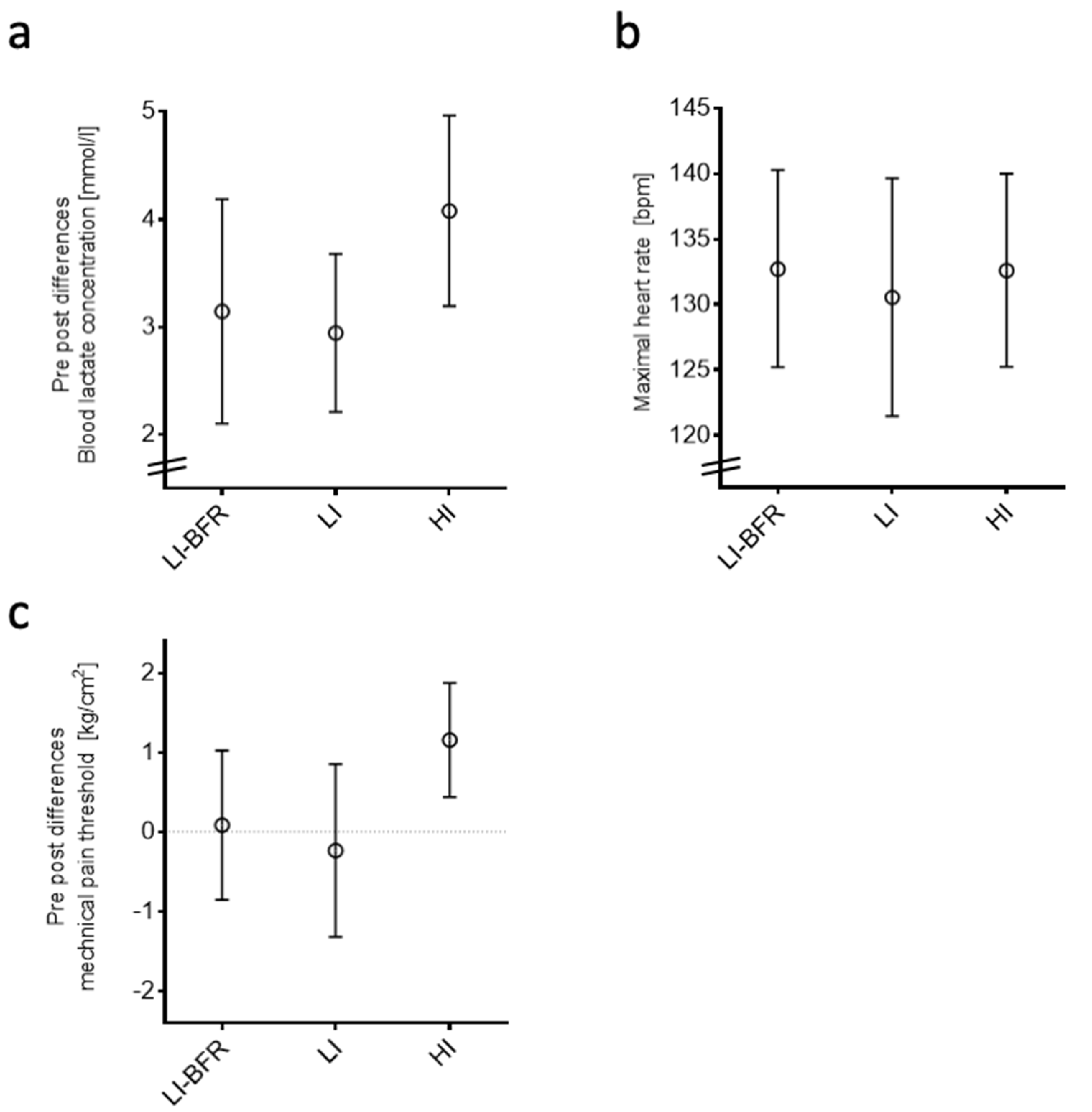
2.3. Basic Resistance Training Outcomes
2.3.1. Objective Outcomes of the Training Interventions
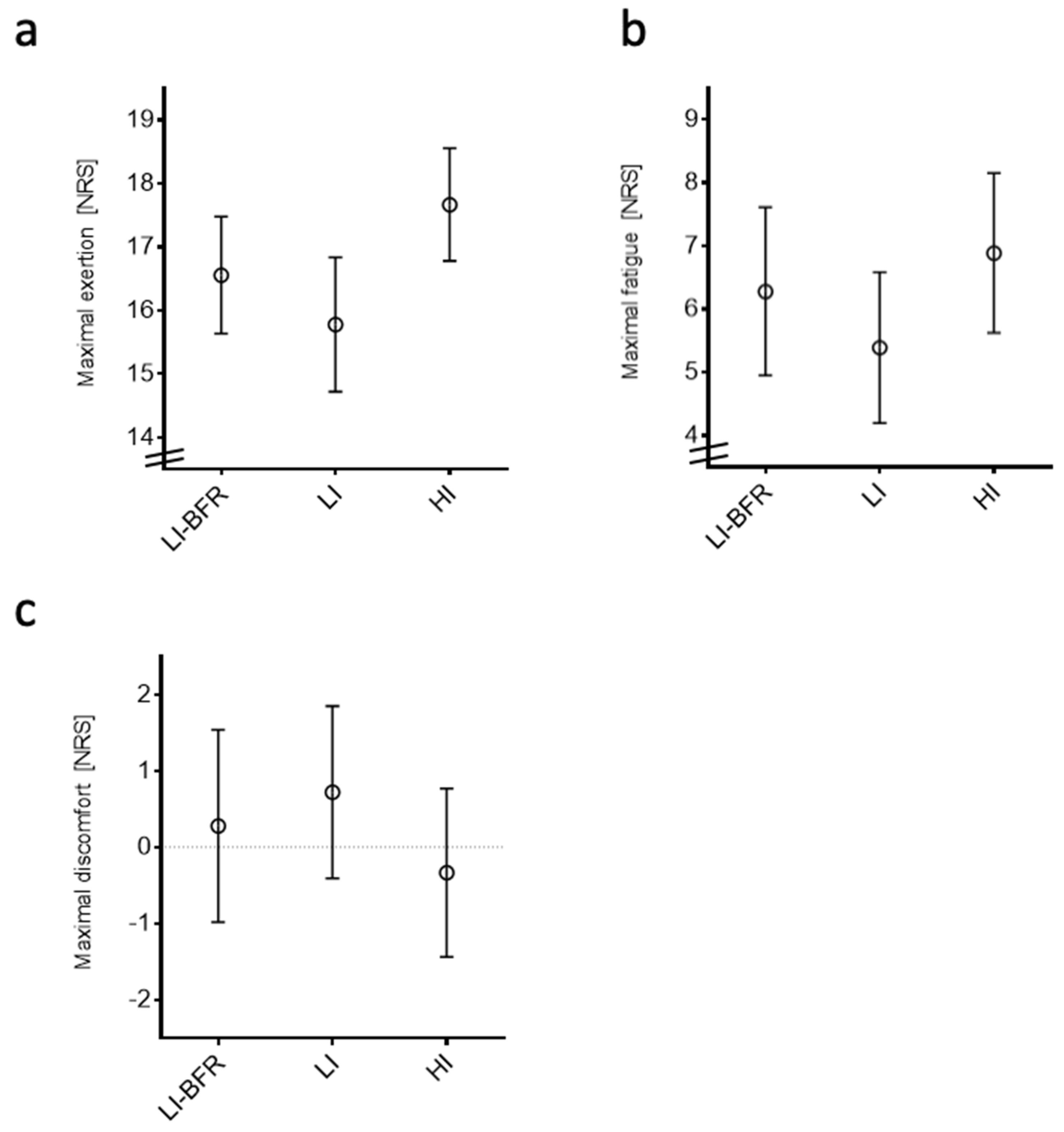
2.3.2. Participant-Reported Outcomes
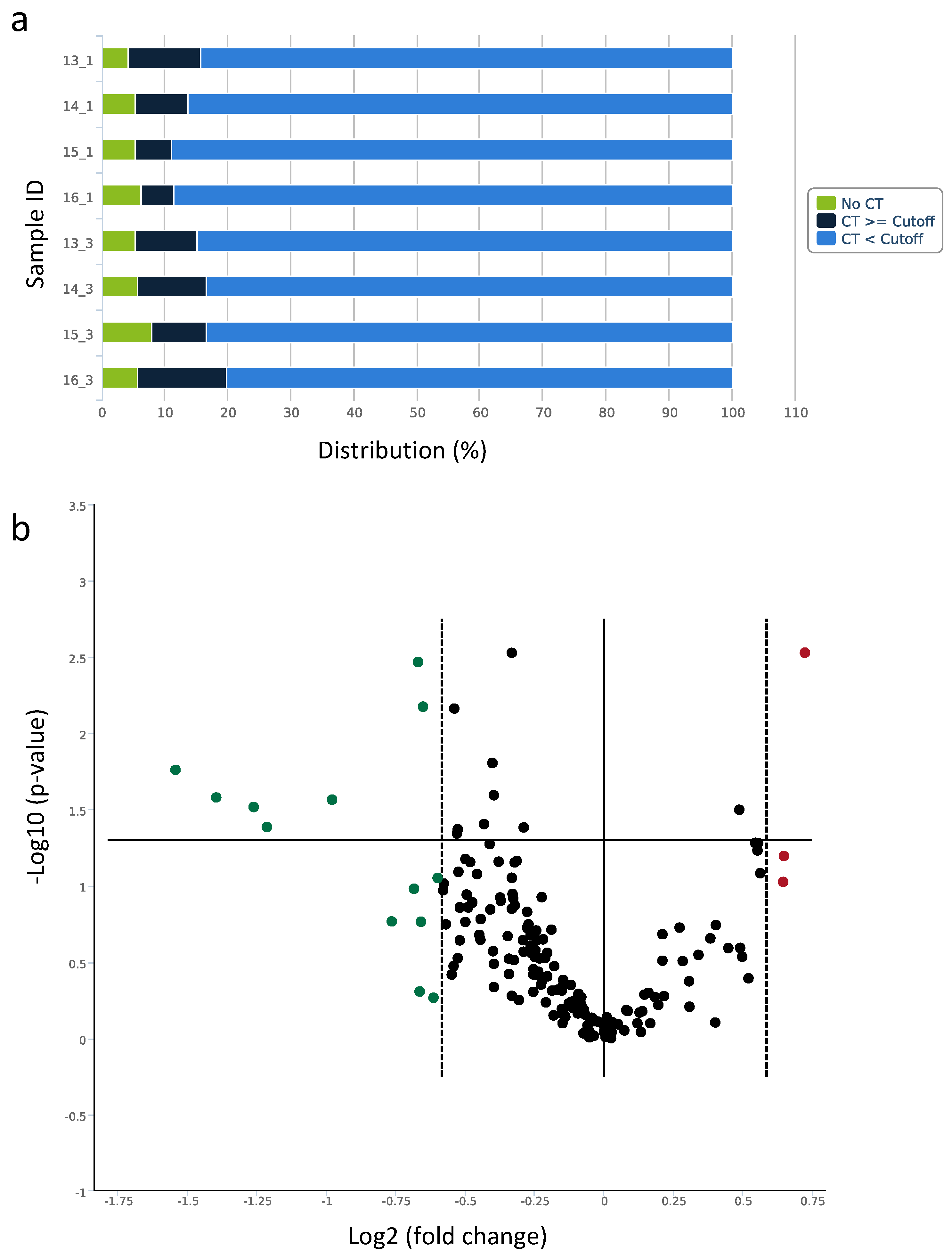
2.4. Profiling of Circulating miRNAs
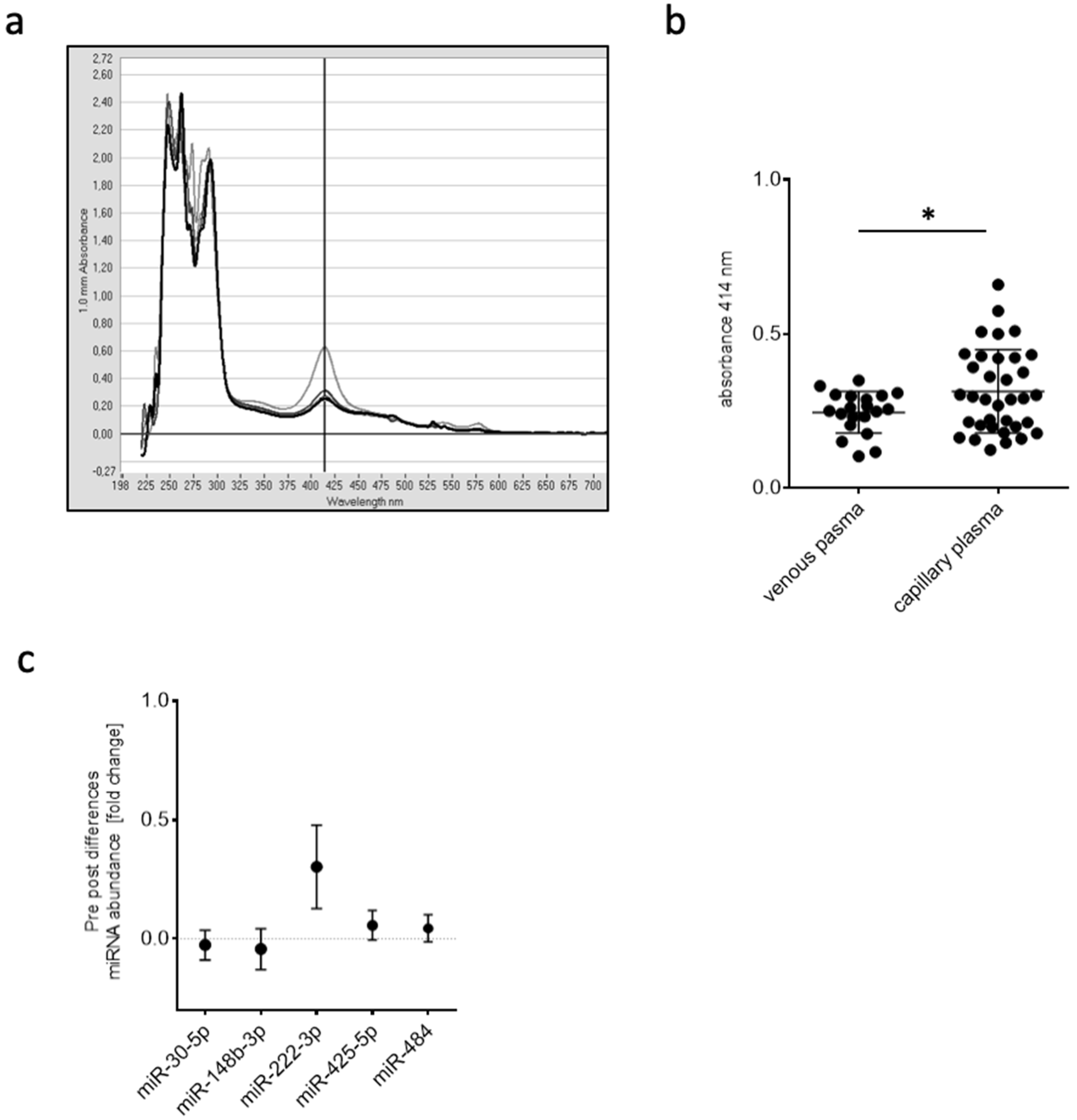
2.4.1. Capillary Blood for miRNA Isolation, Expression Analysis, and Quantification
2.4.2. Screening of Expression Changes in Circulating miRNAs before and after BFR Training
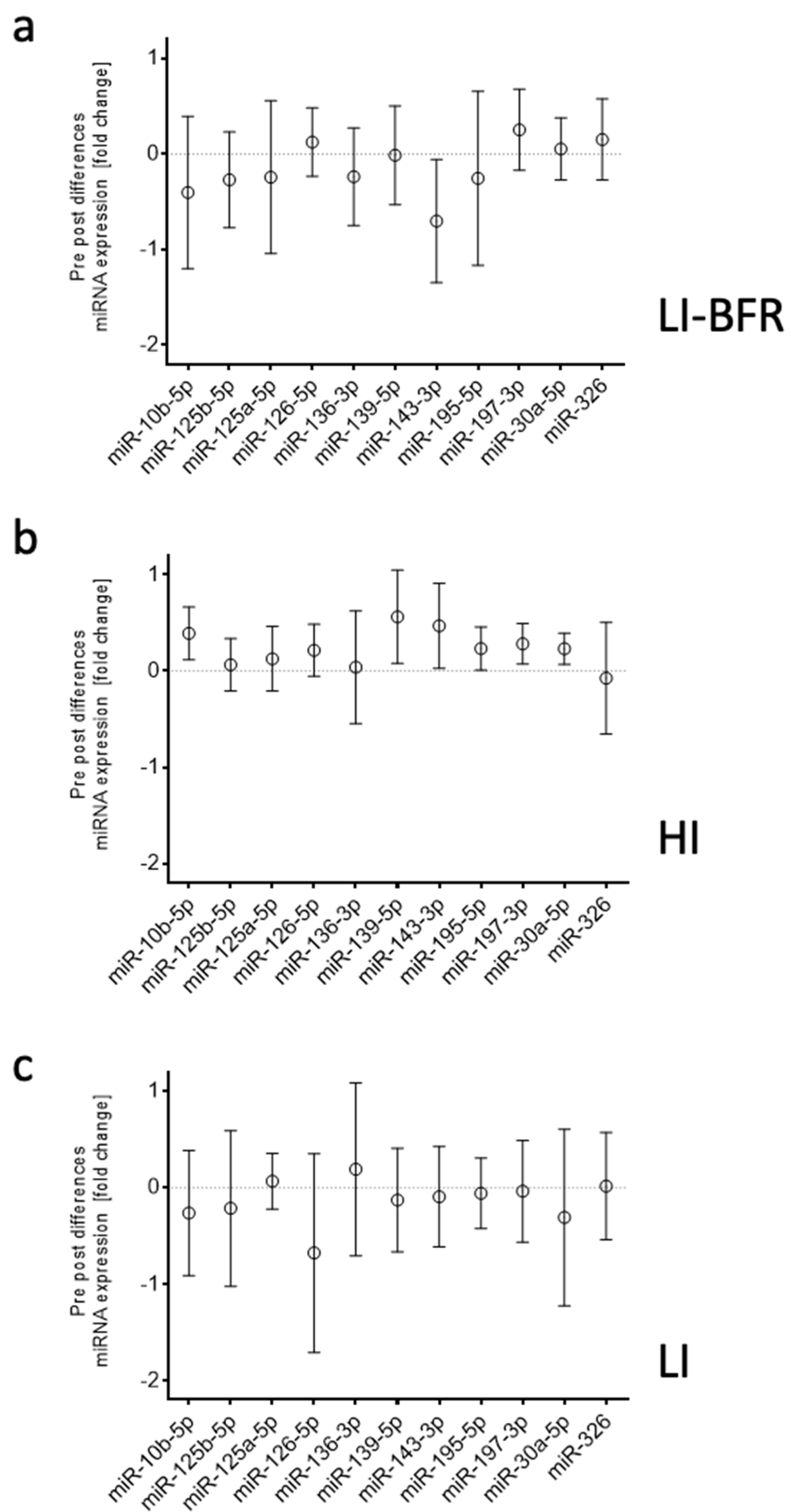
2.4.3. Analysis of miRNAs in Different Training Intervention Groups
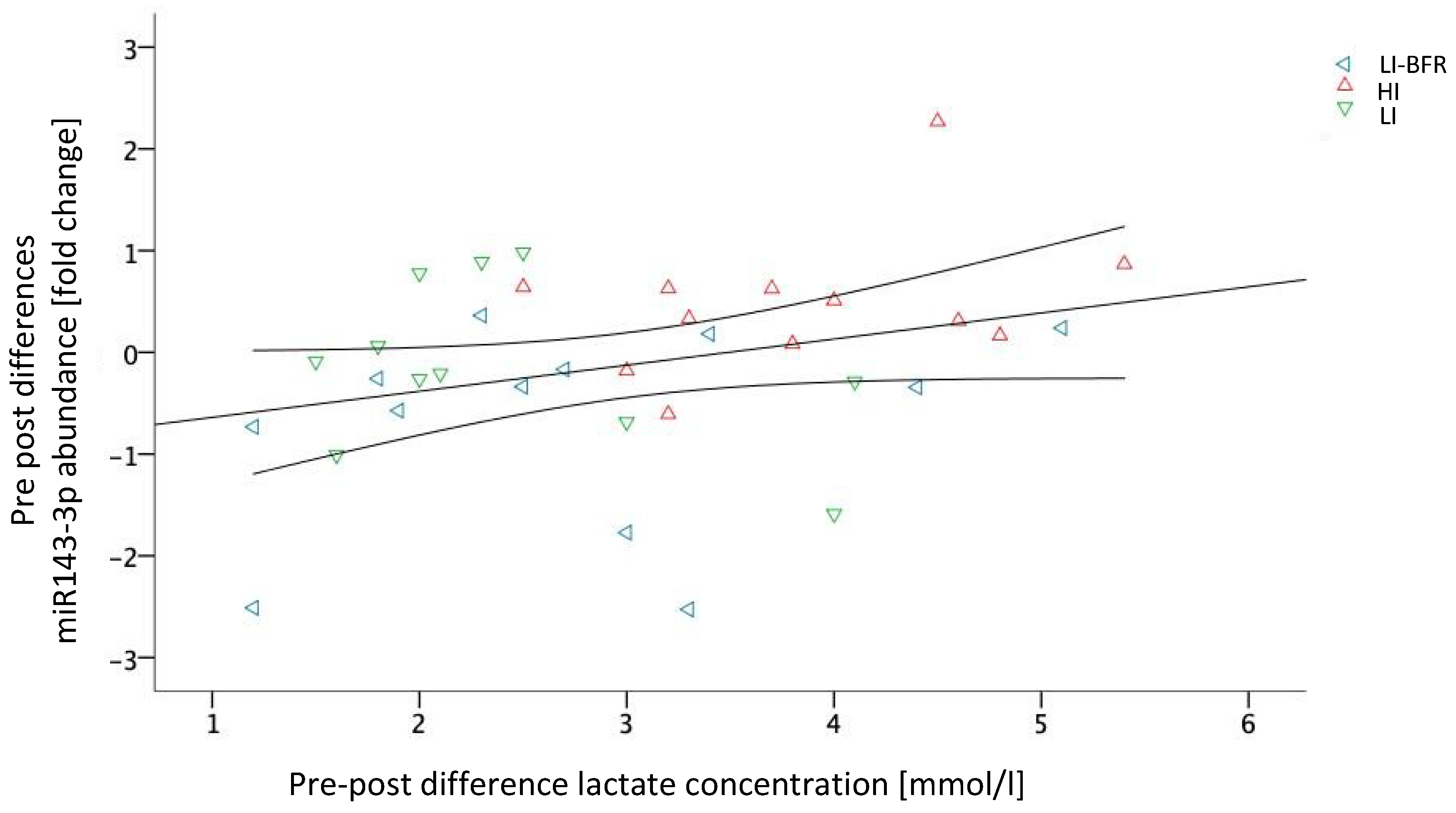
2.5. Associations between Training Outcomes and Circulating miRNAs
3. Discussion
4. Materials and Methods
4.1. Ethical Standard and Study Design
4.2. Sample
4.3. Experimental Design
4.4. Blood Flow Velocity Measurement
4.5. RM Determination
4.6. Intervention
4.7. Assessments
4.7.1. Laboratory Analytic Outcomes
Blood Lactate Concentration
Heart Rate
Mechanical Pain Threshold
Blood Sampling and Plasma Preparation for miRNA Profiling
Determination of Hemolysis
miRNA Isolation
Reverse Transcription and miRNA Profiling
Reverse Transcription and qRT-PCR
4.7.2. Self-Reported Outcomes
4.8. Data Analyses and Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| miRNA | microRNA |
| PAD | Peripheral artery disease |
| BFR | Blood flow restriction |
| FSS | Fluid shear stress |
| HI | High intensity |
| LI | Low intensity |
| LI-BFR | Low intensity with blood flow restriction |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| hsa-miR-30e-5p | YP00204714 | 5′UGUAAACAUCCUUGACUGGAAG |
| hsa-miR-148b-3p | YP00204047 | 5′UCAGUGCAUCACAGAACUUUGU |
| hsa-miR-222-3p | YP00204551 | 5′AGCUACAUCUGGCUACUGGGU |
| hsa-miR-425-5p | YP00204337 | 5′AAUGACACGAUCACUCCCGUUGA |
| hsa-miR-484 | YP00205636 | 5′UCAGGCUCAGUCCCCUCCCGAU |
| hsa-miR-197-3p | YP00204380 | 5′UUCACCACCUUCUCCACCCAGC |
| hsa miR-326 | YP00204512 | 5′CCUCUGGGCCCUUCCUCCAG |
| hsa-miR-136-3p | YP00205503 | 5′CAUCAUCGUCUCAAAUGAGUCU |
| hsa-miR-143-3p | YP00205992 | 5′UGAGAUGAAGCACUGUAGCUC |
| hsa-miR-30a-5p | YP00205695 | 5′UGUAAACAUCCUCGACUGGAAG |
| hsa-miR-139-5p | YP00205874 | 5′UCUACAGUGCACGUGUCUCCAGU |
| hsa-miR-125a-5p | YP00204339 | 5′UCCCUGAGACCCUUUAACCUGUGA |
| hsa-miR-125b-5p | YP00205713 | 5′UCCCUGAGACCCUAACUUGUGA |
| hsa-miR-126-5p | YP00206010 | 5′CAUUAUUACUUUUGGUACGCG |
| hsa-miR-10b-5p | YP00205637 | 5′UACCCUGUAGAACCGAAUUUGUG |
| hsa-miR-195-5p | YP00205869 | 5′UAGCAGCACAGAAAUAUUGGC |
References
- Schaper, W. On arteriogenesis—A reply. Basic Res. Cardiol. 2003, 98, 183–184. [Google Scholar] [PubMed]
- Heil, M.; Eitenmüller, I.; Schmitz-Rixen, T.; Schaper, W. Arteriogenesis versus angiogenesis: Similarities and differences. J. Cell. Mol. Med. 2006, 10, 45–55. [Google Scholar] [CrossRef]
- Ben Driss, A.; Benessiano, J.; Poitevin, P.; Levy, B.I.; Michel, J.B. Arterial expansive remodeling induced by high flow rates. Am. J. Physiol. 1997, 272, H851–H858. [Google Scholar] [CrossRef] [PubMed]
- Gerhold, K.A.; Schwartz, M.A. Ion Channels in Endothelial Responses to Fluid Shear Stress. Physiology 2016, 31, 359–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Z.-D.; Tarbell, J.M. Fluid Flow Mechanotransduction in Vascular Smooth Muscle Cells and Fibroblasts. Ann. Biomed. Eng. 2011, 39, 1608–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tronc, F.; Mallat, Z.; Lehoux, S.; Wassef, M.; Esposito, B.; Tedgui, A. Role of matrix metalloproteinases in blood flow-induced arterial enlargement: Interaction with NO. Arterioscler. Thromb. Vasc. Biol. 2000, 20, E120–E126. [Google Scholar] [CrossRef]
- Neth, P.; Nazari-Jahantigh, M.; Schober, A.; Weber, C. MicroRNAs in flow-dependent vascular remodelling. Cardiovasc. Res. 2013, 99, 294–303. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.-K.; Zhu, J.-Q.; Zhang, J.-T.; Li, Q.; Li, Y.; He, J.; Qin, Y.-W.; Jing, Q. Circulating microRNA: A novel potential biomarker for early diagnosis of acute myocardial infarction in humans. Eur. Heart J. 2010, 31, 659–666. [Google Scholar] [CrossRef]
- Zhang, C. MicroRNAs in Vascular Biology and Vascular Disease. J. Cardiovasc. Trans. Res. 2010, 3, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Chan, S.Y.; Zhang, Y.-Y.; Hemann, C.; Mahoney, C.E.; Zweier, J.L.; Loscalzo, J. MicroRNA-210 Controls Mitochondrial Metabolism during Hypoxia by Repressing the Iron-Sulfur Cluster Assembly Proteins ISCU1/2. Cell Metab. 2009, 10, 273–284. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.H.; Liu, N.; van Rooij, E.; Olson, E.N. MicroRNA control of muscle development and disease. Curr. Opin. Cell Biol. 2009, 21, 461–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hergenreider, E.; Heydt, S.; Tréguer, K.; Boettger, T.; Horrevoets, A.J.G.; Zeiher, A.M.; Scheffer, M.P.; Frangakis, A.S.; Yin, X.; Mayr, M.; et al. Atheroprotective communication between endothelial cells and smooth muscle cells through miRNAs. Nat. Cell Biol. 2012, 14, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-J.; Wei, S.-Y.; Chiu, J.-J. Mechanical regulation of epigenetics in vascular biology and pathobiology. J. Cell. Mol. Med. 2013, 17, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Huonker, M.; Halle, M.; Keul, J. Structural and functional adaptations of the cardiovascular system by training. Int. J. Sports Med. 1996, 17, S164–S172. [Google Scholar] [CrossRef] [PubMed]
- Nash, M.S.; Montalvo, B.M.; Applegate, B. Lower extremity blood flow and responses to occlusion ischemia differ in exercise-trained and sedentary tetraplegic persons. Arch. Phys. Med. Rehab. 1996, 77, 1260–1265. [Google Scholar] [CrossRef]
- Dopheide, J.F.; Rubrech, J.; Trumpp, A.; Geissler, P.; Zeller, G.C.; Schnorbus, B.; Schmidt, F.; Gori, T.; Münzel, T.; Espinola-Klein, C. Supervised exercise training in peripheral arterial disease increases vascular shear stress and profunda femoral artery diameter. Eur. J. Prev. Cardiol. 2017, 24, 178–191. [Google Scholar] [CrossRef]
- Sayed, A.; Schierling, W.; Troidl, K.; Rüding, I.; Nelson, K.; Apfelbeck, H.; Benli, I.; Schaper, W.; Schmitz-Rixen, T. Exercise Linked to Transient Increase in Expression and Activity of Cation Channels in Newly Formed Hind-limb Collaterals. Eur. J. Vasc. Endovasc. Surg. 2010, 40, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.L.; Halperin, J.L.; Albert, N.M.; Bozkurt, B.; Brindis, R.G.; Curtis, L.H.; DeMets, D.; Guyton, R.A.; Hochman, J.S.; Kovacs, R.J.; et al. Management of patients with peripheral artery disease (compilation of 2005 and 2011 ACCF/AHA guideline recommendations): A report of the American College of Cardiology Foundation/American Heart Association Task Force on Practice Guidelines. Circulation 2013, 127, 1425–1443. [Google Scholar] [CrossRef]
- Tendera, M.; Aboyans, V.; Bartelink, M.-L.; Baumgartner, I.; Clément, D.; Collet, J.-P.; Cremonesi, A.; de Carlo, M.; Erbel, R.; Fowkes, F.G.R.; et al. ESC Guidelines on the diagnosis and treatment of peripheral artery diseases: Document covering atherosclerotic disease of extracranial carotid and vertebral, mesenteric, renal, upper and lower extremity arteries: The Task Force on the Diagnosis and Treatment of Peripheral Artery Diseases of the European Society of Cardiology (ESC). Eur. Heart J. 2011, 32, 2851–2906. [Google Scholar]
- Niessner, A.; Richter, B.; Penka, M.; Steiner, S.; Strasser, B.; Ziegler, S.; Heeb-Elze, E.; Zorn, G.; Leitner-Heinschink, A.; Niessner, C.; et al. Endurance training reduces circulating inflammatory markers in persons at risk of coronary events: Impact on plaque stabilization? Atherosclerosis 2006, 186, 160–165. [Google Scholar] [CrossRef]
- Michishita, R.; Shono, N.; Inoue, T.; Tsuruta, T.; Node, K. Effect of exercise therapy on monocyte and neutrophil counts in overweight women. Am. J. Med. Sci. 2010, 339, 152–156. [Google Scholar] [PubMed]
- Timmerman, K.L.; Flynn, M.G.; Coen, P.M.; Markofski, M.M.; Pence, B.D. Exercise training-induced lowering of inflammatory (CD14+CD16+) monocytes: A role in the anti-inflammatory influence of exercise? J. Leuk. Biol. 2008, 84, 1271–1278. [Google Scholar] [CrossRef] [PubMed]
- Higashi, Y.; Sasaki, S.; Kurisu, S.; Yoshimizu, A.; Sasaki, N.; Matsuura, H.; Kajiyama, G.; Oshima, T. Regular aerobic exercise augments endothelium-dependent vascular relaxation in normotensive as well as hypertensive subjects: Role of endothelium-derived nitric oxide. Circulation 1999, 100, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, L.F.; Rocha, A.M.; Martins, C.N.; Ribeiro, J.P.; Wally, C.; Strieder, D.L.; Carissimi, C.G.; Oliveira, M.G.; Pereira, A.A.; Biondi, H.S.; et al. Oxidative status of the myocardium in response to different intensities of physical training. Physiol. Res. 2016, 65, 737–749. [Google Scholar] [PubMed]
- Menêses, A.L.; Ritti-Dias, R.M.; Parmenter, B.; Golledge, J.; Askew, C.D. Combined Lower Limb Revascularisation and Supervised Exercise Training for Patients with Peripheral Arterial Disease: A Systematic Review of Randomised Controlled Trials. Sports Med. 2017, 47, 987–1002. [Google Scholar] [CrossRef] [PubMed]
- Takano, H.; Morita, T.; Iida, H.; Asada, K.; Kato, M.; Uno, K.; Hirose, K.; Matsumoto, A.; Takenaka, K.; Hirata, Y.; et al. Hemodynamic and hormonal responses to a short-term low-intensity resistance exercise with the reduction of muscle blood flow. Eur. J. Appl. Physiol. 2005, 95, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Green, D.J.; Hopman, M.T.E.; Padilla, J.; Laughlin, M.H.; Thijssen, D.H.J. Vascular Adaptation to Exercise in Humans: Role of Hemodynamic Stimuli. Physiol. Rev. 2017, 97, 495–528. [Google Scholar] [CrossRef]
- Loenneke, J.P.; Abe, T.; Wilson, J.M.; Thiebaud, R.S.; Fahs, C.A.; Rossow, L.M.; Bemben, M.G. Blood flow restriction: An evidence based progressive model. Acta Physiol. Hung. 2012, 99, 235–250. [Google Scholar] [CrossRef]
- Alberti, G.; Cavaggioni, L.; Silvaggi, N.; Caumo, A.; Garufi, M. Resistance Training with Blood Flow Restriction Using the Modulation of the Muscle’s Contraction Velocity. Strength Cond. J. 2013, 35, 42–47. [Google Scholar]
- Formenti, D.; Perpetuini, D.; Iodice, P.; Cardone, D.; Michielon, G.; Scurati, R.; Alberti, G.; Merla, A. Effects of knee extension with different speeds of movement on muscle and cerebral oxygenation. PeerJ 2018, 6, e5704. [Google Scholar] [CrossRef] [Green Version]
- Bagley, J.R.; Rosengarten, J.J.; Galpin, A.J. Is Blood Flow Restriction Training Beneficial for Athletes? Strength Cond. J. 2015, 37, 48–53. [Google Scholar] [CrossRef]
- Loenneke, J.P.; Pujol, T.J. The Use of Occlusion Training to Produce Muscle Hypertrophy. Strength Cond. J. 2009, 31, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Cook, S.B.; Brown, K.A.; Deruisseau, K.; Kanaley, J.A.; Ploutz-Snyder, L.L. Skeletal muscle adaptations following blood flow-restricted training during 30 days of muscular unloading. J. Appl. Physiol. 2010, 109, 341–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gualano, B.; Neves, M.; Lima, F.R.; Pinto, A.L.D.S.; Laurentino, G.; Borges, C.; Baptista, L.; Artioli, G.G.; Aoki, M.S.; Moriscot, A.; et al. Resistance training with vascular occlusion in inclusion body myositis: A case study. Med. Sci. Sports Exerc. 2010, 42, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Guirro, E.C.O.; Leite, G.P.M.F.; Dibai-Filho, A.V.; Borges, N.C.S.; Guirro, R.R.J. Intra- and Inter-rater Reliability of Peripheral Arterial Blood Flow Velocity by Means of Doppler Ultrasound. J. Manipul. Physiol. Therap. 2017, 40, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Venous flow volume measured by duplex ultrasound can be used as an indicator of impaired tissue perfusion in patients with peripheral arterial disease. Med. Ultrason 2015, 17.
- Baggish, A.L.; Hale, A.; Weiner, R.B.; Lewis, G.D.; Systrom, D.; Wang, F.; Wang, T.J.; Chan, S.Y. Dynamic regulation of circulating microRNA during acute exhaustive exercise and sustained aerobic exercise training: Circulating microRNA in exercise. J. Physiol. 2011, 589, 3983–3994. [Google Scholar] [CrossRef]
- Troidl, K.; Hammerschick, T.; Albarran-Juarez, J.; Jung, G.; Schierling, W.; Krüger, M.; Matuschke, B.; Triodl, C.; Schaper, W.; Schmitz-Rixen, T.; et al. Shear stress-induced miR-143-3p in collateral arteries contributes to outward vessel growth by targeting collagenV-α2. “Manuscript submitted for publication”.
- Lagos-Quintana, M. Identification of Novel Genes Coding for Small Expressed RNAs. Science 2001, 294, 853–858. [Google Scholar] [CrossRef] [Green Version]
- Tiedt, S.; Prestel, M.; Malik, R.; Schieferdecker, N.; Duering, M.; Kautzky, V.; Stoycheva, I.; Böck, J.; Northoff, B.H.; Klein, M.; et al. RNA-Seq Identifies Circulating miR-125a-5p, miR-125b-5p, and miR-143-3p as Potential Biomarkers for Acute Ischemic Stroke. Circ. Res. 2017, 121, 970–980. [Google Scholar] [CrossRef]
- Margolis, L.M.; Lessard, S.J.; Ezzyat, Y.; Fielding, R.A.; Rivas, D.A. Circulating MicroRNA Are Predictive of Aging and Acute Adaptive Response to Resistance Exercise in Men. GERONA 2016, glw243. [Google Scholar] [CrossRef]
- Barber, J.L.; Zellars, K.N.; Barringhaus, K.G.; Bouchard, C.; Spinale, F.G.; Sarzynski, M.A. The Effects of Regular Exercise on Circulating Cardiovascular-related MicroRNAs. Sci. Rep. 2019, 9, 7527. [Google Scholar] [CrossRef] [PubMed]
- Domańska-Senderowska, D.; Laguette, M.-J.; Jegier, A.; Cięszczyk, P.; September, A.; Brzeziańska-Lasota, E. MicroRNA Profile and Adaptive Response to Exercise Training: A Review. Int. J. Sports Med. 2019, 40, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Karabulut, M.; Abe, T.; Sato, Y.; Bemben, M.G. The effects of low-intensity resistance training with vascular restriction on leg muscle strength in older men. Eur. J. Appl. Physiol. 2010, 108, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Borg, G.A. Psychophysical bases of perceived exertion. Med. Sci. Sports Exerc. 1982, 14, 377–381. [Google Scholar] [CrossRef]







| miRNA ID | Fold Change Screen | p-Value | HI | LI-BFR | LI |
|---|---|---|---|---|---|
| hsa-miR-197-3p | 1.56 | 0.094 | |||
| hsa-miR-326 | 1.65 | 0.002 ** | |||
| hsa-miR-136-3p | 1.57 | 0.063 | |||
| hsa-miR-143-3p | −1.7 | 0.170 | up * | down * | |
| hsa-miR-30a-5p | −1.59 | 0.003 ** | up ** | ||
| hsa-miR-139-5p | −1.97 | 0.027 * | up * | ||
| hsa-miR-125a-5p | −2.63 | 0.026 * | |||
| hsa-miR-375 | −1.59 | 0.492 | |||
| hsa-miR-99a-5p | −1.58 | 0.171 | |||
| hsa-miR-126-5p | −1.57 | 0.006 ** | |||
| hsa-miR-10b-5p | −2.4 | 0.030 * | up ** | ||
| hsa-miR-195-5p | −2.92 | 0.017 * | up * | ||
| hsa-miR-125b-5p | −2.32 | 0.041 * | |||
| hsa-miR-100-5p | −1.61 | 0.104 | |||
| hsa-miR-362-3p | −1.52 | 0.088 | |||
| hsa-miR-376c-3p | −1.53 | 0.540 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vogel, J.; Niederer, D.; Engeroff, T.; Vogt, L.; Troidl, C.; Schmitz-Rixen, T.; Banzer, W.; Troidl, K. Effects on the Profile of Circulating miRNAs after Single Bouts of Resistance Training with and without Blood Flow Restriction—A Three-Arm, Randomized Crossover Trial. Int. J. Mol. Sci. 2019, 20, 3249. https://doi.org/10.3390/ijms20133249
Vogel J, Niederer D, Engeroff T, Vogt L, Troidl C, Schmitz-Rixen T, Banzer W, Troidl K. Effects on the Profile of Circulating miRNAs after Single Bouts of Resistance Training with and without Blood Flow Restriction—A Three-Arm, Randomized Crossover Trial. International Journal of Molecular Sciences. 2019; 20(13):3249. https://doi.org/10.3390/ijms20133249
Chicago/Turabian StyleVogel, Johanna, Daniel Niederer, Tobias Engeroff, Lutz Vogt, Christian Troidl, Thomas Schmitz-Rixen, Winfried Banzer, and Kerstin Troidl. 2019. "Effects on the Profile of Circulating miRNAs after Single Bouts of Resistance Training with and without Blood Flow Restriction—A Three-Arm, Randomized Crossover Trial" International Journal of Molecular Sciences 20, no. 13: 3249. https://doi.org/10.3390/ijms20133249