Distinct and Overlapping Expression Patterns of the Homer Family of Scaffolding Proteins and Their Encoding Genes in Developing Murine Cephalic Tissues


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Specificities of Homer Probes and Anti-Homer Antibodies
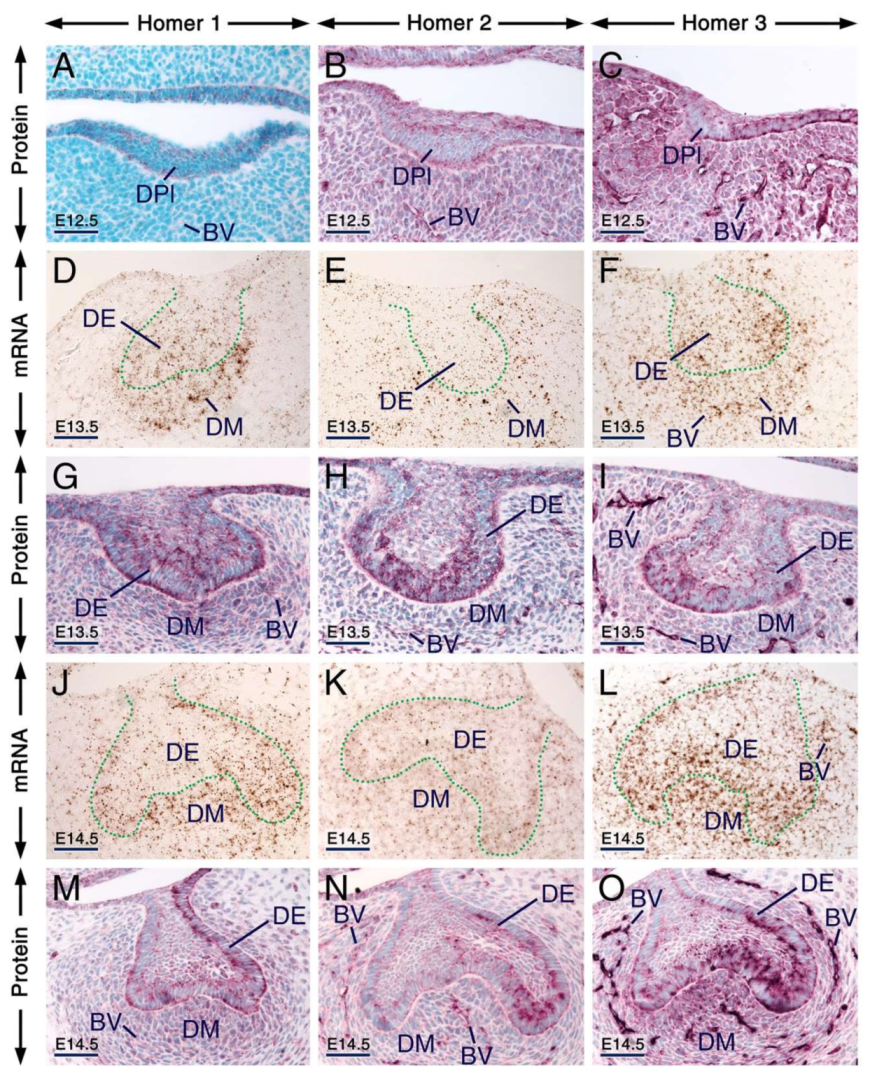
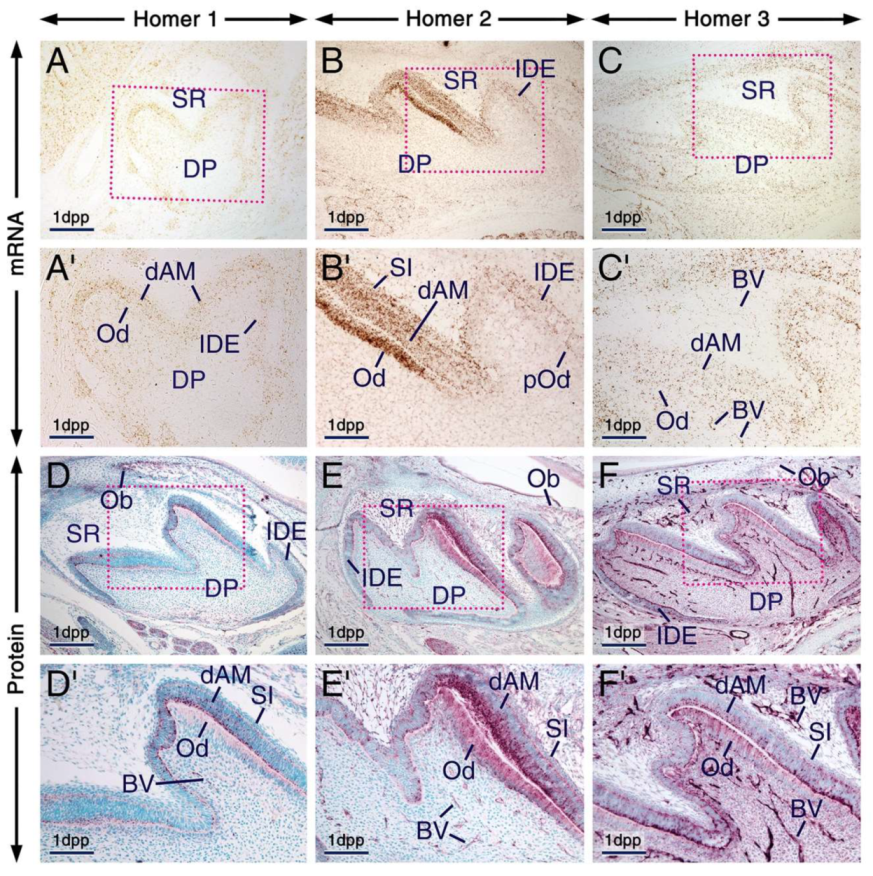
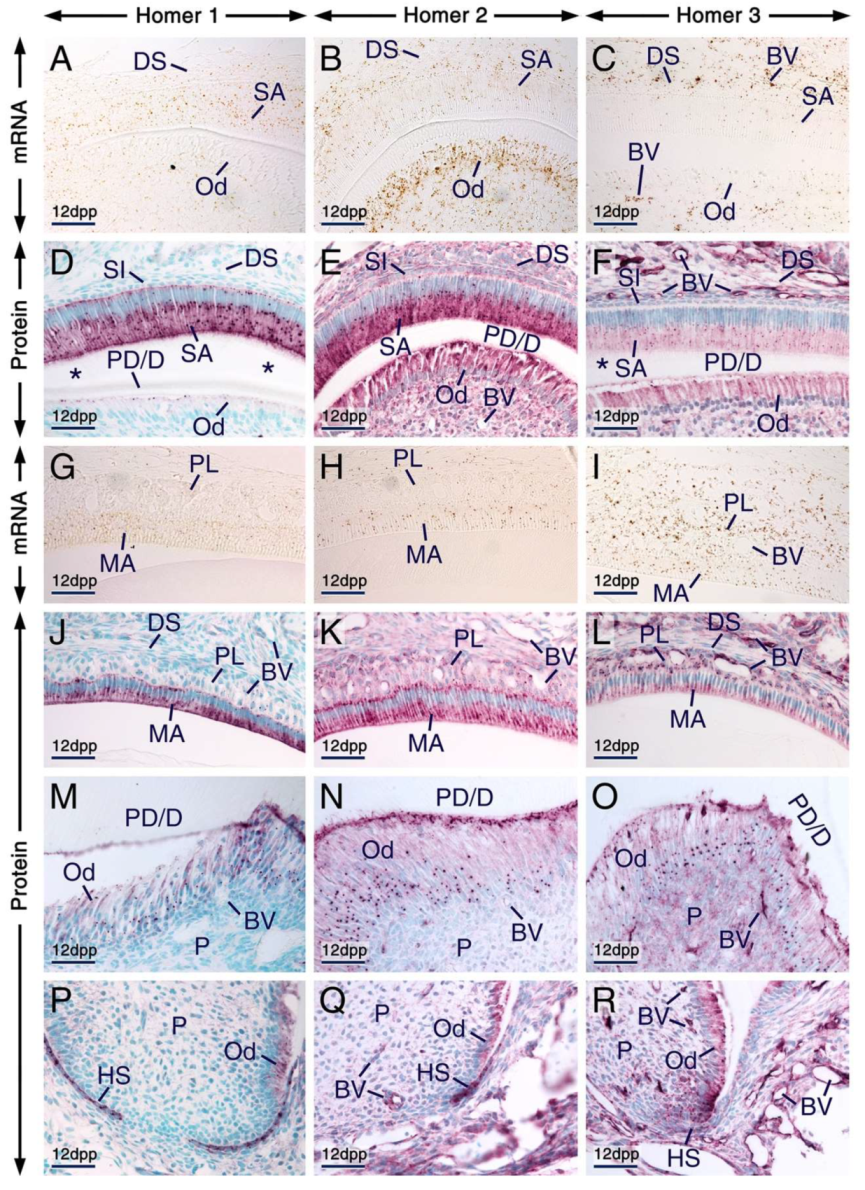
2.2. Expression Patterns of Homer1, Homer2 and Homer3 in the Developing Tooth
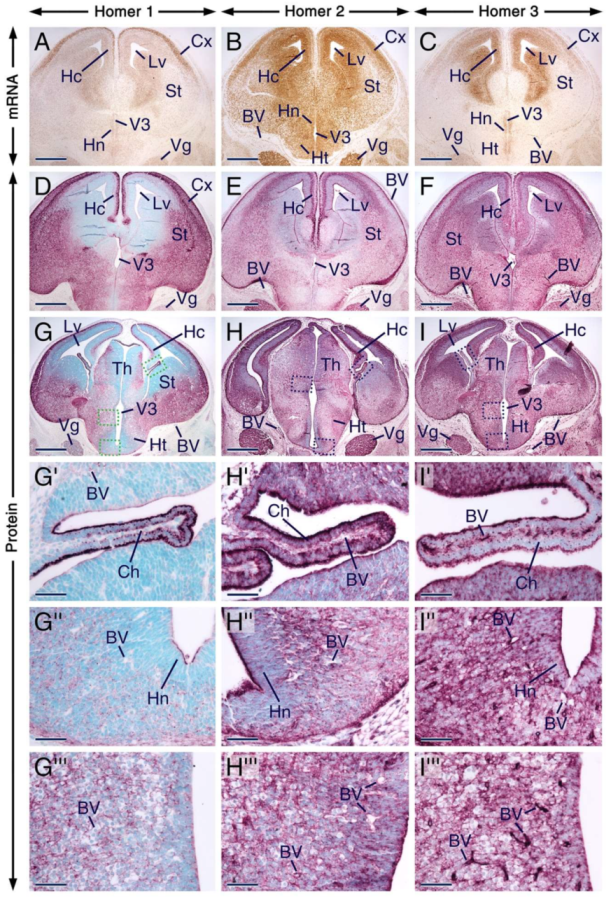
2.3. Expression Patterns of Homer1, Homer2 and Homer3 in the Developing Forebrain and Trigeminal Ganglion
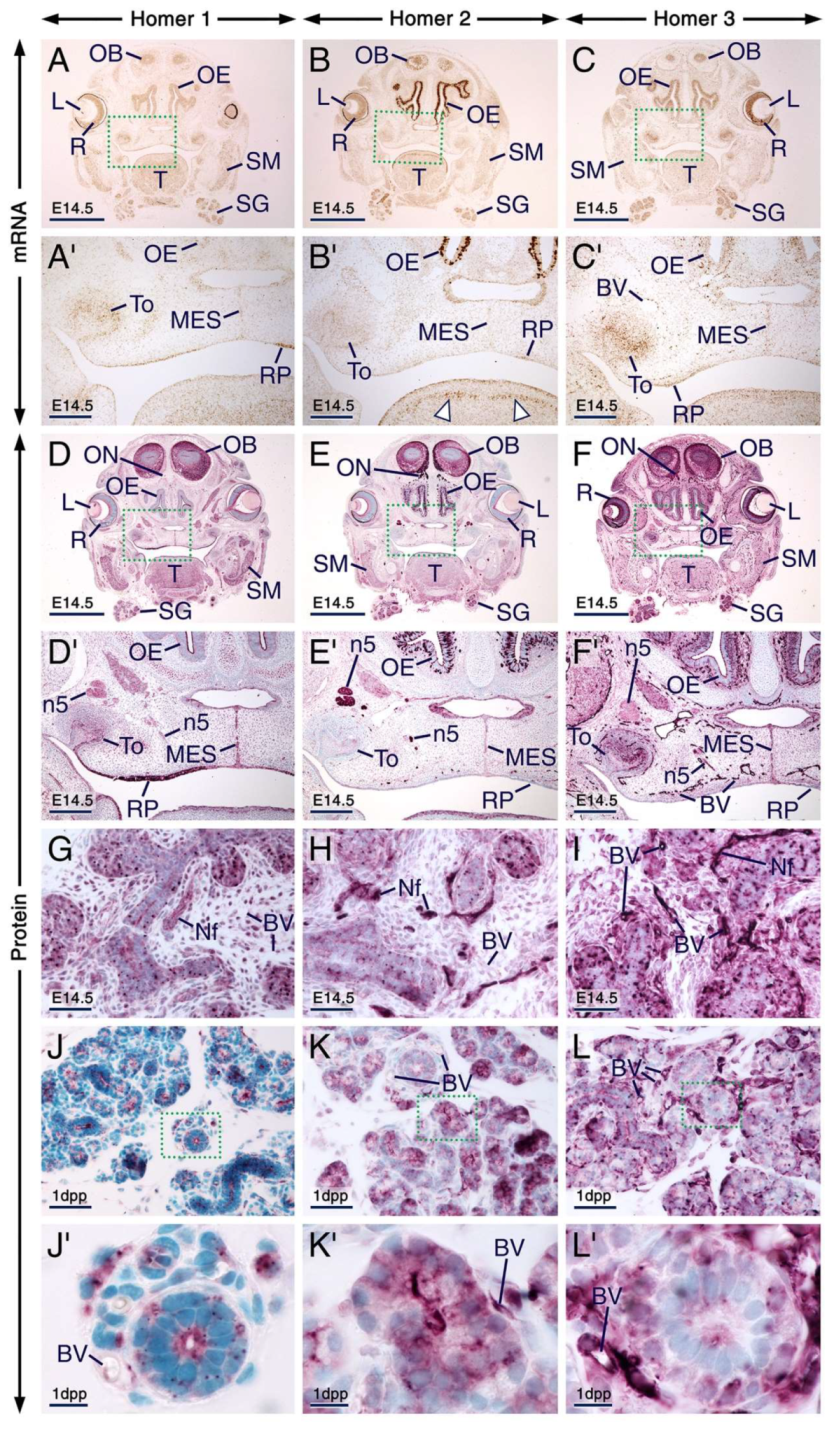
2.4. Expression Patterns of Homer1, Homer2 and Homer 3 in Other Developing Cephalic Regions
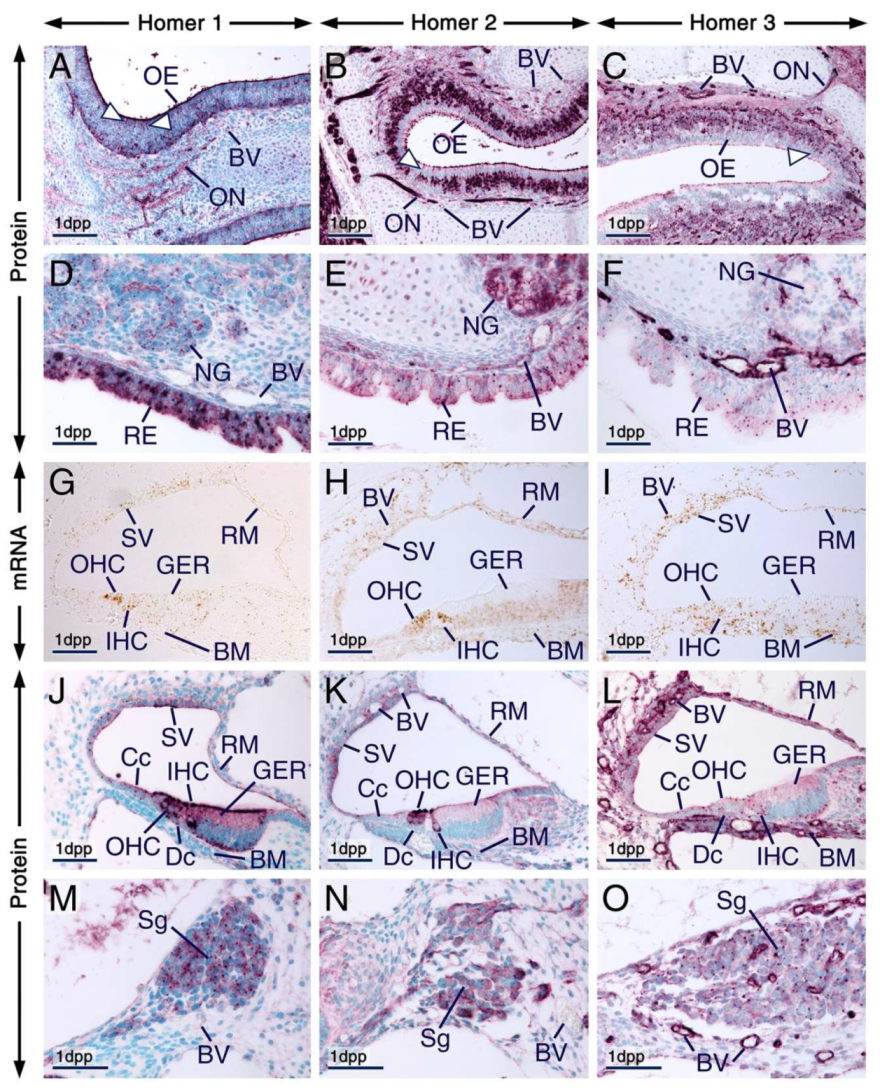
2.4.1. Eye, Olfactory and Respiratory Mucosae, Cranial Nerves, Salivary Glands, Rugae Palatinae, Palatal Medial Epithelial Seam and Skeletal Muscle
2.4.2. Cochlea
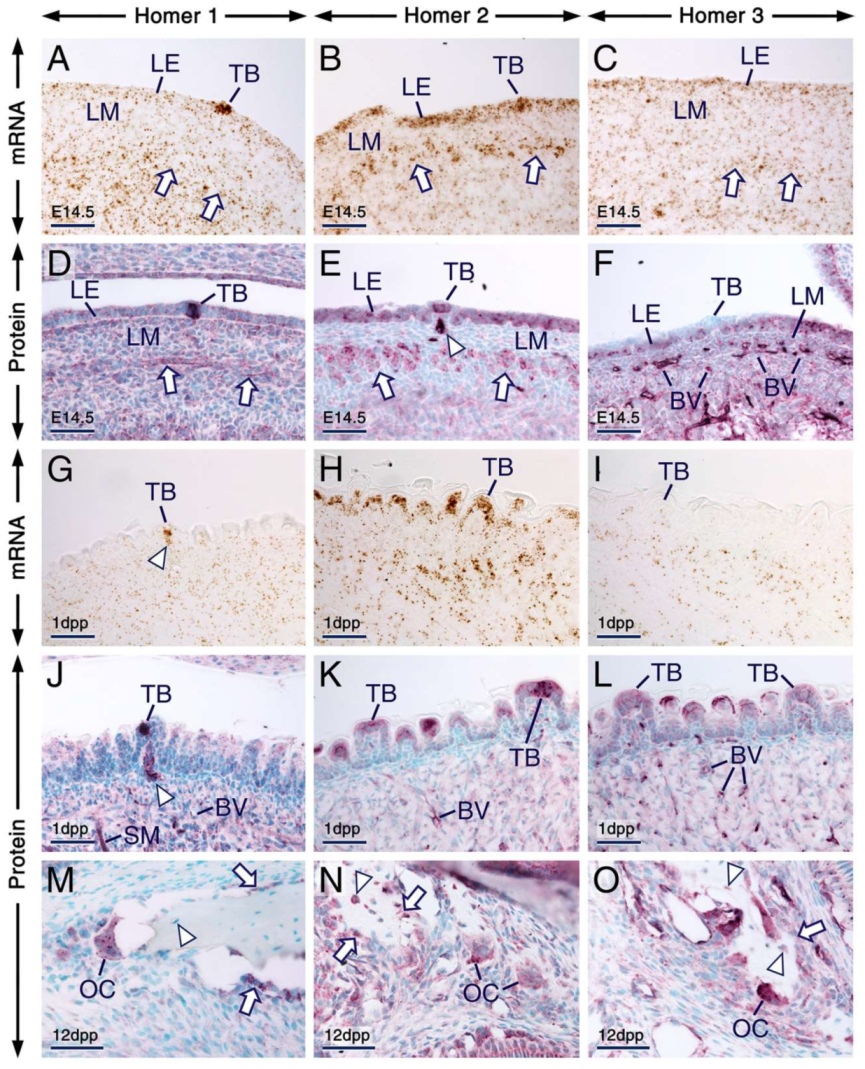
2.4.3. Tongue, Taste Buds and Bone
3. Discussion
3.1. The Three Homer Family Members are Expressed Ubiquitously in Developing Cephalic Tissues
3.2. Probable Roles for Homer Proteins During Tooth Formation
3.3. Overlapping Expression Patterns and Distinct Subcellular Localization of Homer Proteins
3.4. Homer Proteins Are Enriched in Puncta
3.5. The Vascular Endothelium Is Enriched with Homer3 and also Produces Homer1 and Homer2
4. Materials and Methods
4.1. Ethics Statement
4.2. Tissue Processing, Immunohistochemistry and in Situ Hybridization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BSA | Bovine serum albumin |
| GER | Greater epithelial ridge of the developing organ of Corti |
| IHC | Inner hair cells of the organ of Corti |
| IP3Rs | Inositol 1,4,5-triphosphate receptors |
| mGluR1/5 | Group I metabotropic glutamate receptors |
| mTOR | Mammalian/mechanistic target of Rapamycin |
| NFATc | Nuclear factor of activated T-cells |
| OHC | Outer hair cells of the organ of Corti |
| PBS | Phosphate buffered saline |
| PMCA | Plasma-membrane Ca2+-ATPase pump |
| RANKL | Receptor activator of nuclear factor kappa-B ligand |
| RT-qPCR | Reverse transcription quantitative polymerase chain reaction |
| RyRs | Ryanodine receptors |
| SOCE | Store-operated calcium entry |
| STIM1/STIM2 | Stromal interaction molecules 1 and 2 |
| TRPC | Transient receptor potential canonical |
References
- Brakeman, P.R.; Lanahan, A.A.; O’Brien, R.; Roche, K.; Barnes, C.A.; Huganir, R.L.; Worley, P.F. Homer: A protein that selectively binds metabotropic glutamate receptors. Nature 1997, 386, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Ozawa, F.; Saito, Y.; Hirai, K.; Inokuchi, K. Vesl, a gene encoding VASP/Ena family related protein, is upregulated during seizure, long-term potentiation and synaptogenesis. Febs. Lett. 1997, 412, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Xiao, B.; Tu, J.C.; Petralia, R.S.; Yuan, J.P.; Doan, A.; Breder, C.D.; Ruggiero, A.; Lanahan, A.A.; Wenthold, R.J.; Worley, P.F. Homer regulates the association of group I metabotropic glutamate receptors with multivalent complexes of homer-related, synaptic proteins. Neuron 1998, 21, 707–716. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Tadokoro, S.; Imanaka, T.; Murakami, S.D.; Nakamura, M.; Kashiwada, K.; Ko, J.; Nishida, W.; Sobue, K. Isolation of PSD-Zip45, a novel Homer/vesl family protein containing leucine zipper motifs, from rat brain. Febs. Lett. 1998, 437, 304–308. [Google Scholar] [CrossRef] [Green Version]
- Shiraishi, Y.; Mizutani, A.; Bito, H.; Fujisawa, K.; Narumiya, S.; Mikoshiba, K.; Furuichi, T. Cupidin, an isoform of Homer/Vesl, interacts with actin cytoskeleton and activates Rho family of small GTPases and is expressed in developing mouse cerebellar granule cells. J. Neurosci. 1999, 19, 8389–8400. [Google Scholar] [CrossRef] [Green Version]
- Kato, A.; Ozawa, F.; Saitoh, Y.; Fukazawa, Y.; Sugiyama, H.; Inokichi, K. Novel members of the vesl/Homer family of PDZ proteins that bind metabotropic glutamate receptors. J. Biol. Chem. 1998, 273, 23969–23975. [Google Scholar] [CrossRef] [Green Version]
- Shiraishi-Yamaguchi, Y.; Furuichi, T. The Homer family proteins. Genome Biol. 2007, 8, 206. [Google Scholar] [CrossRef] [Green Version]
- Jardin, I.; Lopez, J.J.; Berna-Erro, A.; Rosado, J.A. Homer proteins in Ca2+ entry. IUBMB Life 2013, 65, 497–504. [Google Scholar] [CrossRef]
- Barzik, M.; Carl, U.D.; Schubert, W.D.; Frank, R.; Wehland, J.; Heinz, D.W. The N-terminal domain of Homer/Vesl is a new class II EVH1 domain. J. Mol. Biol. 2001, 309, 155–169. [Google Scholar] [CrossRef]
- Irie, K.; Nakatsu, T.; Mitsuoka, K.; Miyazawa, A.; Sobue, K.; Hiroaki, Y.; Doi, T.; Fujiyoshi, Y.; Kato, H. Crystal structure of the Homer 1 family conserved region reveals the interaction between the EVH1 domain and own proline-rich motif. J. Mol. Biol. 2002, 318, 1117–1126. [Google Scholar] [CrossRef]
- Yuan, J.P.; Kiselyov, K.; Shin, D.M.; Chen, J.; Shcheykinov, N.; Kang, S.H.; Dehoff, M.H.; Schwarrz, M.-K.; Seeburg, P.H.; Muallem, S.; et al. Homer binds TRPC family channels and is required for gating of TRPC1 by IP3 receptors. Cell 2003, 114, 777–789. [Google Scholar] [CrossRef] [Green Version]
- Tu, J.C.; Xiao, B.; Yuan, J.P.; Lanajan, A.A.; Leoffert, K.; Li, M.; Linden, D.J.; Worley, P.F. Homer binds a novel proline-rich motif and links group 1 metabotropic glutamate receptors with IP3 receptors. Neuron 1998, 21, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Duncan, R.S.; Hwang, S.-Y.; Koulen, P. Effects of Vesl/Homer proteins on intracellular signaling. Exp. Biol. Med. 2005, 230, 527–535. [Google Scholar] [CrossRef] [Green Version]
- Worley, P.F.; Zeng, W.; Huang, G.; Kim, J.Y.; Shin, D.M.; Kim, M.S.; Yuan, J.P.; Kiselyov, K.; Muallem, S. Homer proteins in excitable and non-excitable cells. Cell Calcium. 2007, 42, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Tadokoro, S.; Tachibana, T.; Imanaka, T.; Nishida, W.; Sobue, K. Involvement of unique leucine-zipper motif of PSD-Zip45 (Homer1c/vesl-1L) in group I metabotropic glutamate receptor clustering. Proc. Natl. Acad. Sci. USA 1999, 96, 13801–13806. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, M.K.; Ames, H.M.; Hayashi, Y. Tetrameric hub structure of postsynaptic scaffolding protein homer. J. Neurosci. 2006, 26, 8492–8501. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, M.K.; Tang, C.; Verpelli, C.; Narayanan, R.; Stearns, M.H.; Xu, R.-M.; Li, H.; Sala, C.; Hayashi, Y. The postsynaptic density proteins Homer and Shank form a polymeric network structure. Cell 2009, 137, 159–171. [Google Scholar] [CrossRef] [Green Version]
- Shiraishi, Y.; Mizutani, A.; Yuasa, S.; Mikoshiba, K.; Furuichi, T. Differential expression of Homer family proteins in the developing mouse brain. J. Comp. Neurol. 2004, 473, 582–599. [Google Scholar] [CrossRef]
- Li, J.; Zhang, W.; Yang, H.; Howrigan, D.P.; Wilkinson, B.; Souaiaia, T.; Abel, T.; Knowles, J.A.; Clementel, V.A.; Tudor, J.C.; et al. Spatio-temporal profile of postsynaptic interactomes integrates components of complex brain disorders. Nat. Neurosci. 2017, 20, 1150–1161. [Google Scholar] [CrossRef]
- Foa, L.; Indrani, R.; Haas, K.; Wu, G.-Y.; Brakeman, P.; Worley, P.; Cline, H. The scaffold protein, Homer1b/c, regulates axon pathfinding in the central nervous system in vivo. Nat. Neurosci. 2001, 4, 499–506. [Google Scholar] [CrossRef]
- Tanaka, M.; Scott Duncan, R.; McClung, N.; Yannazzo, J.A.; Hwang, S.-Y.; Marunouchi, T.; Inokuchi, K.; Koulen, P. Homer proteins control neuronal differentiation through IP3 receptor signaling. Febs. Lett. 2006, 580, 6145–6150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szumlinski, K.K.; Dehoff, M.H.; Kang, S.H.; Frys, K.A.; Lominac, K.D.; Kligmann, M.; Rohrer, J.; Griffin III, W.; Toda, S.; Champtiaux, N.P.; et al. Homer proteins regulate sensitivity to cocaine. Neuron 2004, 43, 401–413. [Google Scholar] [CrossRef] [Green Version]
- Szumlinski, K.K.; Lominac, K.D.; Oleson, E.B.; Walker, J.K.; Mason, A.; Dehoff, M.H.; Klugmann, M.; Cagle, S.; Welt, K.; During, M.; et al. Homer 2 is necessary for EtOH-induced neuroplasticity. J. Neurosci. 2005, 25, 7054–7061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, K.V.; Häusl, A.; Pöhlmann, M.L.; Hartmann, J.; Labermaier, C.; Muller, M.; Scmidt, M.V. Hippocampal Homer1 levels influence motivational behavior in an operant conditioning task. PLoS ONE 2014, 9, e85975. [Google Scholar] [CrossRef] [PubMed]
- Norton, N.; Williams, H.J.; Williams, N.M.; Spurlock, G.; Zammit, S.; Jones, G.; Jones, S.; Owen, R.; O’Donovan, M.C.; Owen, M.J. Mutation screening of the Homer gene family and association analysis in schizophrenia. Am. J. Med. Genet. B Neuropsychiatr Genet. 2003, 120B, 18–21. [Google Scholar] [CrossRef] [PubMed]
- Uys, J.D.; LaLumiere, R.T. Glutamate: The new frontier in pharmacotherapy for cocaine addition. CNS Neurol. Disord. Drug Targets 2008, 7, 482–491. [Google Scholar] [CrossRef]
- Rietschel, M.; Mattheisen, M.; Frank, J.; Treutlein, J.; Degenhardt, F.; Breuer, R.; Steffens, M.; Mier, D.; Esslinger, C.; Walter, H.; et al. Genome-wide association-, replication-, and neuroimaging study implicates HOMER1 in the etiology of major depression. Biol. Psychiatry 2010, 68, 578–585. [Google Scholar] [CrossRef]
- Sandona, D.; Tibaldo, E.; Volpe, P. Evidence for the presence of two Homer 1 transcripts in skeletal and cardiac muscles. Biochem. Biophys. Res. Commun. 2000, 279, 348–353. [Google Scholar] [CrossRef]
- Soloviev, M.M.; Ciruella, F.; Chan, W.Y.; McIIhinney, R.A. Mouse brain and muscle tissues constitutively express Homer proteins. Eur. J. Biochem. 2000, 267, 634–639. [Google Scholar] [CrossRef] [Green Version]
- Salanova, M.; Bortoloso, E.; Schiff, G.; Gustmann, M.; Belavy, D.L.; Felsenberg, D.; Furlan, S.; Volpe, P.; Blottner, D. Expression and regulation of Homer in human skeletal muscle during neuromuscular juction adaptation to disuse and excercice. FASEB J. 2011, 25, 4312–4325. [Google Scholar] [CrossRef]
- Salanova, M.; Volpe, P.; Blottner, D. Homer protein family regulation in skeletal muscle and neuromuscular adaptation. IUBMB Life 2013, 65, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.M.; Dehoff, M.; Luo, X.; Kang, S.H.; Tu, J.; Nayak, S.K.; Ross, E.M.; Worley, P.F.; Muallem, S. Homer2 tunes G protein-coupled receptors stimulus intensity by regulating RGS proteins and PLC GAP activity. J. Cell Biol. 2003, 162, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-M.; Lee, J.; Jo, H.; Park, S.; Chang, I.; Muallem, S.; Shin, D.M. Homer2 protein regulates plasma membrane Ca2+-ATPase-mediated Ca2+ signaling in mouse parotid gland acinar cells. J. Biol. Chem. 2014, 289, 24971–24979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiber, J.A.; Zhang, Z.-S.; Burch, J.; Eu, J.P.; Zhang, S.; Truskey, G.A.; Seth, M.; Yamaguchi, N.; Meissner, G.; Shah, R.; et al. Mice lacking Homer 1 exhibit a skeletal myopathy characterized by abnormal transient receptor potential channel activity. Mol. Cell Biol. 2008, 28, 2637–2647. [Google Scholar] [CrossRef] [Green Version]
- Azaiez, H.; Decker, A.R.; Booth, K.T.; Simpson, A.C.; Shearer, A.E.; Huygen, P.L.; Bu, F.; Hildebrand, M.S.; Ranum, P.T.; Shibata, S.B.; et al. Homer2, a stereociliary scaffolding protein, is essential for normal hearing in humans and mice. PLoS Genet. 2015, 11, e105137. [Google Scholar] [CrossRef] [Green Version]
- Bavencoffe, A.; Zhu, M.X.; Tian, J.B. New aspects of the contribution of ER to SOCE regulation: TRPC proteins as a link between plasma membrane ion transport and intracellular Ca2+ stores. Adv. Exp. Med. Biol. 2017, 993, 239–255. [Google Scholar]
- Kurnellas, M.P.; Lee, A.K.; Li, H.; Deng, L.; Ehrlich, D.J.; Elkabes, S. Molecular alterations in the cerebellum of the plasma membrane calcium ATPase2 (PMCA2)-null mouse indicate abnormalities in Purkinje neurons. Mol. Cell Neurosci. 2007, 34, 178–188. [Google Scholar] [CrossRef] [Green Version]
- Salm, E.J.; Thayer, S.A. Homer proteins accelerate Ca2+ clearence mediated by the plasma membrane Ca2+ pump in hippocampal neurons. Biochem. Biophys. Res. Commun. 2012, 424, 76–81. [Google Scholar] [CrossRef] [Green Version]
- Jardin, I.; Albarrán, L.; Bermejo, N.; Salido, G.M.; Rosado, J.A. Homers regulate calcium entry and aggregation in human platelets: A role for Homers in the association between STIM1 and Orai1. Biochem. J. 2012, 445, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Jia, S.; Rodriguez, M.; Williams, A.G.; Yuan, J.P. Homer binds to Orai1 and TRPC channels in the neointima and regulate smooth muscle cell migration and proliferation. Sci. Rep. 2017, 7, 5075. [Google Scholar] [CrossRef] [Green Version]
- Prakrya, M.; Lewis, R.S. Store-operated calcium channels. Physiol. Rev. 2015, 95, 1383–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, S.Y.; Wen, X.; Yin, K.; Smith, C.E.; Paine, M.L. Multiple calcium exchangers and pumps are a prominent feature of enamel organ cells. Front. Physiol 2017, 8, 336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Choi, M.; Richardson, A.S.; Reid, B.M.; Seymen, F.; Yildirim, M.; Tuna, E.; Gencay, K.; Simmer, J.P.; Hu, J.C. STIM1 and SLC24A4 are critical for enamel maturation. J. Dent. Res. 2014, 93, 94S–100S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurbaeva, M.K.; Eckstein, M.; Axel, R.C.; Smith, C.E.; Srikanth, C.; Paine, M.L.; Gwack, Y.; Hubbard, M.J.; Feske, S.; Lacruz, R.S. Dental enamel cells express functional SOCE channels. Sci. Rep. 2015, 30, 15803. [Google Scholar] [CrossRef] [Green Version]
- Nurbaeva, M.K.; Eckstein, M.; Feske, S.; Lacruz, R.S. Ca2+ transport and signaling in enamel cells. J. Physiol. 2017, 595, 3015–3039. [Google Scholar] [CrossRef] [Green Version]
- Eckstein, M.; Vaeth, M.; Fornai, C.; Vinu, M.; Bromage, T.G.; Nurbaeva, M.K.; Sorge, J.L.; Coelho, P.G.; Idaghdour, Y.; Feske, S.; et al. Store-operated Ca2+ entry controls ameloblast cell function and enamel development. J. CI Insight. 2017, 2, e91166. [Google Scholar] [CrossRef] [Green Version]
- Eckstein, M.; Vaeth, M.; Aulestia, F.J.; Costiniti, V.; Kassam, S.N.; Bromage, T.G.; Pedersen, P.; Issekutz, T.; Idaghdour, Y.; Moursi, A.M.; et al. Differential regulation of Ca2+ influx by ORAI channels mediates enamel mineralization. Sci. Signal. 2019, 12. [Google Scholar] [CrossRef]
- Lacruz, R.S.; Feske, S. Diseases caused by mutations of ORAI1 and STIM1. Ann. NY Acad. Sci. 2015, 1356, 45–79. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, Y.; Haruyama, N.; Nakaido, M.; Nakanishi, M.; Ryu, N.; Oh-Hora, M.; Kuremoto, K.; Yoshizaki, K.; Takano, Y.; Takahashi, I. Stim1 regulates enamel mineralization and ameloblast modulation. J. Dent. Res. 2017, 96, 1422–1429. [Google Scholar] [CrossRef]
- Jussila, M.; Thesleff, I. Signaling networks regulating tooth organogenesis and regeneration, and the specification of dental mesenchymal and epithelial lineages. Cold Spring Harb. Perspect. Biol. 2012, 4, a008425. [Google Scholar] [CrossRef]
- Lacruz, R.S.; Habelitz, S.; Wright, T.; Paine, M.L. Dental enamel formation and implications for oral health and disease. Physiol. Rev. 2017, 97, 939–993. [Google Scholar] [CrossRef]
- Gritli-Linde, A.; Bei, M.; Maas, R.; Zhang, X.M.; Linde, A.; McMahon, A.P. Shh signaling within the dental epithelium is necessary for cell proliferation, growth and polarization. Development 2002, 129, 5323–5337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanci, A. Ten Cate’s Oral Histology, 6th ed.; Mosby Inc.: St. Louis, MO, USA, 2003; pp. 192–239. [Google Scholar]
- Ruch, J.V.; Lesot, H.; Bégue-Kirn, C. Odontoblast differentiation. Int. J. Dev. Biol. 1995, 39, 51–68. [Google Scholar]
- Warshawsky, H.; Smith, C.E. Morphological classification of rat incisor ameloblasts. Anat. Rec. 1974, 179, 423–446. [Google Scholar] [CrossRef]
- Garant, P.R.; Nalbandian, J. The fine structure of the papillary region of the mouse enamel organ. Arch. Oral. Biol. 1968, 13, 1167–1185. [Google Scholar] [CrossRef]
- Diez-Roux, G.; Banfi, S.; Sultan, M.; Geffers, L.; Anand, S.; Rozado, D.; Magen, A.; Canidio, E.; Pagani, M.; Peluso, I.; et al. A high-resolution anatomical atlas of the transcriptome in the mouse embryo. PLoS Biol. 2011, 9, e1000582. [Google Scholar] [CrossRef] [PubMed]
- Stiber, J.A.; Tabatabaei, N.; Hawkins, A.F.; Hawke, T.; Worley, P.F.; Williams, R.S.; Rosenberg, P. Homer modulates NFAT-dependent signaling during muscle differentiation. Dev. Biol. 2005, 287, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Uraih, L.; Maronpot, R.R. Normal histology of the nasal cavity and application of special techniques. Env. Health Perspecives 1990, 85, 187–208. [Google Scholar]
- Anniko, M. Embryogenesis of the mammalian inner ear. III. Formation of the tectorial membrane of the CBA/CBA mouse in vivo and in vitro. Anat Embryol. 1980, 160, 301–313. [Google Scholar] [CrossRef]
- El Shahawy, M.; Reibring, C.-G.; Neben, C.L.; Hallberg, K.; Marangoni, P.; Harfe, B.D.; Klein, O.D.; Linde, A.; Gritli-Linde, A. Cell fate specification in the lingual epithelium is controlled by antagonistic activities of Sonic hedgehog and retinoic acid. PLoS Genet. 2017, 13, e1006914. [Google Scholar] [CrossRef] [Green Version]
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The versality and universality of calcium signaling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, B. Receptors and transduction in taste. Nature 2001, 413, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, A.; Kuroda, Y.; Futatsugi, A.; Furuichi, T.; Mikoshiba, K. Phosphorylation of Homer3 by calcium/calmodulin-dependent kinase II regulates a coupling state of its target molecules in Purkinje neurons. J. Neurosci. 2008, 28, 5369–5382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, G.N.; Huso, D.L.; Bouyain, S.; Tu, J.; McCorkell, K.A.; May, M.J.; Zhu, Y.; Lutz, M.; Collins, S.; Dehoff, M.; et al. NFAT binding and regulation of T cell activation by the cytoplasmic scaffolding Homer proteins. Science 2008, 318, 476–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rybchyn, M.S.; Islam, K.S.; Brennan-Speranza, T.C.; Cheng, Z.; Brennan, S.C.; Chang, W.; Mason, R.S.; Conigrave, A.D. Homer1 mediates CaSR-dependent activation of mTOR complex 2 and initiates a novel pathway for AKT-dependent-catenin stabilization in osteoblasts. J. Biol. Chem. 2019, 294, 16337–16350. [Google Scholar] [CrossRef] [PubMed]
- Son, A.; Kang, N.; Kim, K.W.; Yang, Y.M.; Shin, D.M. Homer2 and Homer3 modulate RANKL-induced NFATc1 signaling in osteoclastogenesis and bone metabolism. J. Endocrinol. 2019, 242, 241–249. [Google Scholar] [CrossRef]
- Wu, J.; Pipathsouk, A.; Keizer-Gunnink, A.; Fusetti, F.; Alkema, W.; Liu, S.; Altschuler, S.; Wu, L.; Kortholt, A.; Weiner, O.D. Homer3 regulates the establishment of neutrophil polarity. Mol. Biol. Cell 2015, 26, 1629–1639. [Google Scholar] [CrossRef]
- Bezanilla, M.; Gladfelter, A.S.; Kovar, D.R.; Lee, W.-L. Cytoskeletal dynamics: A view from the membrane. J. Cell Biol. 2015, 209, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Svitkina, T. The actin cytoskeleton and actin-based motility. Cold Spring Harb. Perspect. Biol. 2018, 10, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Khaitlina, S.Y. Intracellular transport based on actin polymerization. Biochemistry (Moscow) 2014, 79, 917–927. [Google Scholar] [CrossRef]
- Galletta, B.J.; Cooper, J.A. Actin and endocytosis: Mechanisms and phylogeny. Curr. Opin. Cell Biol. 2009, 21, 20–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadopulos, A.; Tomatis, V.M.; Kasula, R.; Meunier, F.A. The cortical acto-myosin network: From diffusion barrier to functional gateway in the transport of neurosecretory vesicles to the plasma membrane. Front. Endocrinol. 2013, 4, 153. [Google Scholar] [CrossRef] [Green Version]
- Schreiner, D.; Müller, K.; Werner Hofer, H. The intracellular domain of the human protocadherin hFat1 interacts with Homer signaling scaffolding proteins. Febs. Lett. 2006, 580, 5295–5300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiraishi-Yamaguchi, Y.; Sato, Y.; Sakai, R.; Mizutani, A.; Knopfel, T.; Mori, N.; Mikoshiba, K.; Furuishi, T. Interaction of Cupidin/Homer2 with two actin cytoskeletal regulators. BMC Neurosci. 2009, 10, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nepliouev, I.; Zhang, Z.S.; Stiber, J.A. Effect of oxidative stress on Homer scaffolding proteins. PLoS ONE 2011, 6, e26128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-C.; Guo, Y.-F.; He, H.; Lin, X.; Wang, X.-F.; Zhou, R.; Li, W.-T.; Pam, D.-Y.; Shen, J.; Deng, H.-W. Integrative analysis of genomics and transcriptome data to identify potential functional genes of BMDs in females. J. Bone Min. Res. 2016, 31, 1041–1049. [Google Scholar] [CrossRef] [Green Version]
- Ehrengruber, M.U.; Kato, A.; Inokuchi, K.; Hennou, S. Homer/Vesl proteins and their roles in CNS neurons. Mol. Neurobiol. 2004, 29, 213–227. [Google Scholar] [CrossRef]
- Clifton, N.E.; Trent, S.; Thomas, K.L.; Hall, J. Regulation and function of activity-dependent Homer in Synaptic plasticity. Mol. Neuropsychiatry 2019, 5, 147–161. [Google Scholar] [CrossRef]
- Sgambato-faure, V.; Xiong, Y.; Berke, J.D.; Hyman, S.E.; Strehler, E.E. The Homer-1 protein Ania-3 interacts with the plasma membrane calcium pump. Biochem. Biophys. Res. Commun. 2006, 343, 630–637. [Google Scholar] [CrossRef] [Green Version]
- Linde, A.; Lundgren, T. From serum to the mineral phase. The role of the odontoblast in calcium transport and mineral formation. Int. J. Dev. Biol. 1995, 39, 213–222. [Google Scholar]
- Lundgren, T.; Linde, A. Na+/Ca2+ antiport in membranes of rat incisor odontoblasts. J. Oral. Pathol. 1988, 17, 560–563. [Google Scholar] [CrossRef] [PubMed]
- Lundgren, T.; Linde, A. Voltage-gated calcium channels and non-voltage gated calcium uptake pathways in the rat incisor odontoblast plasma membrane. Calcif. Tissue Int. 1997, 60, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Lundquist, P.; Lundgren, T.; Gritli-Linde, A.; Linde, A. Na+/Ca2+ exchanger isoforms of rat odontoblasts and osteoblasts. Calcif. Tissue Int. 2000, 67, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Shibukawa, Y.; Suzuki, T. Ca2+ signaling mediated by IP3-dependent Ca2+ releasing and store operated Ca2+ channels in rat odontoblasts. J. Bone Min. Res. 2003, 18, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Kim, Y.J.; Paik, S.K.; Cho, Y.S.; Kwon, T.G.; Ahn, D.K.; Kim, S.K.; Yoshida, A.; Bae, Y.C. Expression of metabotropic glutamate receptor mGluR5 in human dental pulp. J. Endod. 2009, 35, 690–694. [Google Scholar] [CrossRef] [PubMed]
- Tsumura, M.; Okumura, R.; Tatsuyama, S.; Ichikawa, H.; Muramatsu, T.; Matsuda, T.; Baba, A.; Suzuki, K.; Kajiya, H.; Sahara, Y.; et al. Ca2+ extrusion via Na+- Ca2+ exchangers in rat odontoblasts. J. Endod. 2010, 36, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Tsumura, M.; Sobhan, U.; Muramatsu, T.; Sato, M.; Ichikawa, H.; Sahara, Y.; Tazaki, M.; Shibukawa, Y. TRPV1-mediated calcium signal couples with cannabinoid receptors and sodium-calcium exchangers in rat odontoblasts. Cell Calcium. 2012, 52, 124–136. [Google Scholar] [CrossRef]
- Nishiyama, A.; Sato, M.; Kimura, M.; Katakura, A.; Tazaki, M.; Shibukawa, Y. Intercellular signal communication among odontoblasts and trigeminal ganglion neurons via glutamate. Cell Calcium. 2016, 60, 341–355. [Google Scholar] [CrossRef]
- Kimura, M.; Nishi, K.; Higashikawa, A.; Ohyama, S.; Sakurai, K.; Tazaki, M.; Shibukawa, Y. High pH-sensitive store-operated Ca2+ entry mediated by Ca2+ release-activated Ca2+ channels in rat odontoblasts. Front. Physiol. 2018, 9, 443. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Zinn, V.; Lefkelidou, A.; Taqi, N.; Chatzistavrou, X.; Balam, T.; Nervina, J.; Papagerakis, S.; Papagerakis, P. Orai1 expression pattern in tooth and craniofacial ectodermal tissues and potential functions during ameloblast differentiation. Dev. Dyn. 2015, 244, 1249–1258. [Google Scholar] [CrossRef] [Green Version]
- Lundgren, T.; Linde, A. Modulation of rat incisor odontoblast plasma membrane-associated Ca2+ with nifedipine. Biochem. Biophys. Acta 1998, 1373, 341–346. [Google Scholar] [CrossRef] [Green Version]
- Westenbroek, E.E.; Anderson, N.L.; Byers, M.R. Altered localization of Cav1.2 (L-type) calcium channels in nerve fibers, Schwann cells, odontoblasts, and fibroblasts of tooth pulp after tooth injury. J. Neurosci. Res. 2004, 75, 371–383. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Deng, X.; Mancarella, S.; Hendron, E.; Eguchi, S.; Soboloff, J.; Tang, X.D.; Gill, D.L. The calcium store sensor, STIM1, reciprocally controls Orai and Cav1.2 channels. Science 2010, 330, 105–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dionisio, N.; Smani, T.; Woodard, G.E.; Castellano, A.; Salido, G.M.; Rosado, J.A. Homer proteins mediate the interactions between STIM1 and Cav1.2 channels. Biochim. Biophysy. Acta 2015, 1853, 1145–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Zhang, Y.; Wang, H.; Zeng, Y.-Y.; Li, Z.; Li, M.-L.; Li, F.-F.; You, J.; Zhang, Z.-M.; Tzeng, C.-M. WW domain-binding protein 2 acts as an oncogene by modulating the activity of the glycolytic enzyme ENO1 in glioma. Cell Death Dis. 2018, 9, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buniello, A.; Ingham, N.J.; Lewis, M.A.; Huma, A.C.; Martinez-Vega, R.; Varela-Nieto, I.; Vizcay-Barrena, G.; Fleck, R.A.; Houston, O.; Bardhan, T.; et al. Wbp2 is required for normal glutamatergic synapses in the cochlea and is crucial for hearing. EMBO Mol. Med. 2016, 8, 191–207. [Google Scholar] [CrossRef]
- Sala, C.; Piech, V.; Wilson, N.R.; Passafaro, M.; Liu, G.; Sheng, M. Regulation of dendritic spine morphology and synaptic function by Shank and Homer. Neuron 2001, 31, 115–130. [Google Scholar] [CrossRef] [Green Version]
- Diagana, T.T.; Thomas, U.; Prokopenko, S.N.; Xiao, B.; Worley, P.F.; Thomas, J.B. Mutation of Drosophila homer disrupts control of locomotor activity and behavioral plasticity. J. Neurosci. 2002, 22, 428–436. [Google Scholar] [CrossRef] [Green Version]
- Smani, T.; Gómez, L.J.; Regodon, S.; Woodard, G.E.; Siegfried, G.; Khatib, A.-M.; Rosado, J.A. TRP channels in angiogenesis and other endothelial functions. Front. Physiol. 2018, 9, 1731. [Google Scholar] [CrossRef]
- Yuan, J.P.; Lee, K.P.; Hong, J.H.; Muallem, S. The closing and opening of TRPC channels by Homer1 and Stim1. Acta Physiol. (Oxf.) 2012, 204, 238–247. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Zeng, W.; Kiselyov, K.; Yuan, J.P.; Dehoff, M.H.; Mikoshiba, K.; Worley, P.F.; Muallem, S. Homer1 mediates store- and inositol 1,4,5-triphosphate receptor-dependent translocation and retrieval of TRPC3 to the plasma membrane. J. Biol. Chem. 2006, 281, 32540–32549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirao, T.; Sekino, Y. General introduction to Drebrin. Adv. Exp. Med. Biol. 2017, 1006, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Reibring, C.-G.; El Shahawy, M.; Hallberg, K.; Kannius-Janson, M.; Nilsson, J.; Parkkila, S.; Sly, W.S.; Waheed, A.; Linde, A.; Gritli-Linde, A. Expression patterns and subcellular localization of carbonic anhydrases are developmentally regulated during tooth formation. PLoS ONE 2014, 9, e96007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gritli-Linde, A.; Lewis, P.; McMahon, A.P.; Linde, A. The whereabouts of a morphogen: Direct evidence for short- and graded long-range activity of Hedgehog signaling peptides. Dev. Biol. 2001, 236, 364–386. [Google Scholar] [CrossRef]
- Wang, F.; Flanagan, J.; Su, N.; Wang, L.-C.; Bui, S.; Nielson, A.; Wu, X.; Vo, H.-T.; Ma, X.-J.; Luo, Y. RNAscope. A Novel in situ RNA analysis platform for formalin-fixed, paraffin-embedded tissues. J. Mol. Diagn. 2012, 14, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Kuboki, Y.; Schatz, C.A.; Koechert, K.; Schubert, S.; Feng, J.; Wittemer-Rump, S.; Ziegelbauer, K.; Krahn, T.; Nagatsuma, A.K.; Ochiai, A. In situ analysis of FGFR2 mRNA and comparison with FGFR2 gene copy number by dual-color in situ hybridization in a large cohort of gastric cancer patients. Gastric. Cancer 2018, 21, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Mead, T.J.; Apte, S.S. Expression analysis by RNAscopeTM in situ hybridization. Methods Mol. Biol. 2020, 2043, 173–178. [Google Scholar]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reibring, C.-G.; Hallberg, K.; Linde, A.; Gritli-Linde, A. Distinct and Overlapping Expression Patterns of the Homer Family of Scaffolding Proteins and Their Encoding Genes in Developing Murine Cephalic Tissues. Int. J. Mol. Sci. 2020, 21, 1264. https://doi.org/10.3390/ijms21041264
Reibring C-G, Hallberg K, Linde A, Gritli-Linde A. Distinct and Overlapping Expression Patterns of the Homer Family of Scaffolding Proteins and Their Encoding Genes in Developing Murine Cephalic Tissues. International Journal of Molecular Sciences. 2020; 21(4):1264. https://doi.org/10.3390/ijms21041264
Chicago/Turabian StyleReibring, Claes-Göran, Kristina Hallberg, Anders Linde, and Amel Gritli-Linde. 2020. "Distinct and Overlapping Expression Patterns of the Homer Family of Scaffolding Proteins and Their Encoding Genes in Developing Murine Cephalic Tissues" International Journal of Molecular Sciences 21, no. 4: 1264. https://doi.org/10.3390/ijms21041264