Molecular Mechanisms, Epidemiology, and Clinical Importance of β-Lactam Resistance in Enterobacteriaceae


 and
and
Abstract
:1. Introduction
2. β-Lactam Molecular Resistance in Enterobacteriaceae: β-Lactamases
2.1. Extended-Spectrum β-Lactamases
2.2. Carbapenemases
2.3. Extended-Spectrum AmpC Cephalosporinases
3. β-Lactamase-Mediated Resistance in Enterobacteriaceae: Epidemiology
3.1. Historical Overview
3.2. Molecular Overview
4. β-Lactamase-Mediated Resistance in Enterobacteriaceae: Clinical Importance
5. Conclusions
Funding
Conflicts of Interest
References
- Abraham, E.P.; Chain, E. An enzyme from bacteria able to destroy penicillin. 1940. Rev. Infect. Dis. 1988, 10, 677–678. [Google Scholar]
- Bush, K.; Bradford, P.A. β-Lactams and β-Lactamase Inhibitors: An Overview. Cold Spring Harb. Perspect. Med. 2016, 6, a025247. [Google Scholar] [CrossRef]
- Waxman, D.J.; Strominger, J.L. Penicillin-binding proteins and the mechanism of action of beta-lactam antibiotics. Annu. Rev. Biochem. 1983, 52, 825–869. [Google Scholar] [CrossRef]
- Meroueh, S.O.; Minasov, G.; Lee, W.; Shoichet, B.K.; Mobashery, S. Structural aspects for evolution of beta-lactamases from penicillin-binding proteins. J. Am. Chem. Soc. 2003, 125, 9612–9618. [Google Scholar] [CrossRef]
- Klein, E.Y.; Van Boeckel, T.P.; Martinez, E.M.; Pant, S.; Gandra, S.; Levin, S.A.; Goossens, H.; Laxminarayan, R. Global increase and geographic convergence in antibiotic consumption between 2000 and 2015. Proc. Natl. Acad. Sci. USA 2018, 115, E3463–E3470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tooke, C.L.; Hinchliffe, P.; Bragginton, E.C.; Colenso, C.K.; Hirvonen, V.H.A.; Takebayashi, Y.; Spencer, J. β-Lactamases and β-Lactamase Inhibitors in the 21st Century. J. Mol. Biol. 2019, 431, 3472–3500. [Google Scholar] [CrossRef] [PubMed]
- Bush, K. Proliferation and significance of clinically relevant β-lactamases. Ann. N. Y. Acad. Sci. 2013, 1277, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Ambler, R.P. The structure of B-lactamases. Philos. Trans. R. Soc. B Biol. Sci. 1980, 289, 321–331. [Google Scholar]
- Rawlings, N.D.; Barrett, A.J.; Bateman, A. MEROPS: The peptidase database. Nucleic Acids Res. 2010, 38, D227–D233. [Google Scholar] [CrossRef] [PubMed]
- Bush, K. Past and Present Perspectives on β-Lactamases. Antimicrob. Agents Chemother. 2018, 62, e01076-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palzkill, T. Structural and Mechanistic Basis for Extended-Spectrum Drug-Resistance Mutations in Altering the Specificity of TEM, CTX-M, and KPC β-lactamases. Front. Mol. Biosci. 2018, 5, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naas, T.; Oueslati, S.; Bonnin, R.A.; Dabos, M.L.; Zavala, A.; Dortet, L.; Retailleau, P.; Iorga, B.I. Beta-lactamase database (BLDB)—Structure and function. J. Enzym. Inhib. Med. Chem. 2017, 32, 917–919. [Google Scholar] [CrossRef] [PubMed]
- Wilson, H.; Török, M.E. Extended-spectrum b-lactamase-producing and carbapenemase-producing Enterobacteriaceae. Microb. Genom. 2018, 4, e000197. [Google Scholar] [CrossRef] [PubMed]
- Friedman, N.D.; Temkin, E.; Carmeli, Y. The negative impact of antibiotic resistance. Clin. Microbiol. Infect. 2016, 22, 416–422. [Google Scholar] [CrossRef]
- Gould, I.M. Antibiotic resistance: The perfect storm. Int. J. Antimicrob. Agents 2009, 34, S2–S5. [Google Scholar] [CrossRef]
- The Review on Antimicrobial Resistance, Chaired by Jim O’Neill. Antimicrobial Resistance: Tackling a Crisis for the Health and Wealth of Nations. December 2014. Available online: http://www.jpiamr.eu/wp-content/uploads/2014/12/AMR-Review-Paper-Tackling-a-crisis-for-the-health-and-wealth-of-nations_1-2.pdf (accessed on 10 July 2020).
- World Health Organization. Global Priority List of Antibiotic—Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; World Health Organization: Geneva, Switzerland, 2017; Available online: https://www.who.int/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf?ua=1 (accessed on 14 February 2020).
- Suay-García, B.; Pérez-Gracia, M.T. Present and Future of Carbapenem-resistant Enterobacteriaceae (CRE) Infections. Antibiotics 2019, 8, 122. [Google Scholar] [CrossRef] [Green Version]
- Vivas, R.; Barbosa, A.A.T.; Dolabela, S.S.; Jain, S. Multidrug-Resistant Bacteria and Alternative Methods to Control Them: An Overview. Microb. Drug Resist. 2019, 25, 890–908. [Google Scholar] [CrossRef]
- Blair, J.M.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef]
- Bonomo, R.A. β-Lactamases: A Focus on Current Challenges. Cold Spring Harb. Perspect. Med. 2017, 7, a025239. [Google Scholar] [CrossRef]
- Rodríguez-Baño, J.; Gutiérrez-Gutiérrez, B.; Machuca, I.; Pascual, A. Treatment of Infections Caused by Extended-Spectrum-Beta-Lactamase-, AmpC-, and Carbapenemase-Producing Enterobacteriaceae. Clin. Microbiol. Rev. 2018, 31, e00079-17. [Google Scholar] [CrossRef] [Green Version]
- Bush, K.; Jacoby, G.A. Updated functional classification of beta-lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walther-Rasmussen, J.; Høiby, N. OXA-type carbapenemases. J. Antimicrob. Chemother. 2006, 57, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liakopoulos, A.; Mevius, D.; Ceccarelli, D. A Review of SHV Extended-Spectrum β-Lactamases: Neglected Yet Ubiquitous. Front. Microbiol. 2016, 7, 1374. [Google Scholar] [CrossRef] [PubMed]
- Knothe, H.; Shah, P.; Krcmery, V.; Antal, M.; Mitsuhashi, S. Transferable resistance to cefotaxime, cefoxitin, cefamandole and cefuroxime in clinical isolates of Klebsiella pneumoniae and Serratia marcescens. Infection 1983, 11, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Sougakoff, W.; Goussard, S.; Gerbaud, G.; Courvalin, P. Plasmid-mediated resistance to third-generation cephalosporins caused by point mutations in TEM-type penicillinase genes. Rev. Infect. Dis. 1988, 10, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Cantón, R.; Coque, T.M. The CTX-M beta-lactamase pandemic. Curr. Opin. Microbiol. 2006, 9, 466–475. [Google Scholar] [CrossRef]
- Bevan, E.R.; Jones, A.M.; Hawkey, P.M. Global epidemiology of CTX-M β-lactamases: Temporal and geographical shifts in genotype. J. Antimicrob. Chemother. 2017, 72, 2145–2155. [Google Scholar] [CrossRef] [Green Version]
- Ramadan, A.A.; Abdelaziz, N.A.; Amin, M.A.; Aziz, R.K. Novel blaCTX-M variants and genotype-phenotype correlations among clinical isolates of extended spectrum beta lactamase-producing Escherichia coli. Sci. Rep. 2019, 9, 4224. [Google Scholar] [CrossRef]
- Cantón, R.; González-Alba, J.M.; Galán, J.C. CTX-M Enzymes: Origin and Diffusion. Front. Microbiol. 2012, 3, 110. [Google Scholar] [CrossRef] [Green Version]
- Branger, C.; Ledda, A.; Billard-Pomares, T.; Doublet, B.; Fouteau, S.; Barbe, V.; Roche, D.; Cruveiller, S.; Médigue, C.; Castellanos, M.; et al. Extended-spectrum β-lactamase-encoding genes are spreading on a wide range of Escherichia coli plasmids existing prior to the use of third-generation cephalosporins. Microb. Genom. 2018, 4, e000203. [Google Scholar] [CrossRef]
- Naas, T.; Poirel, L.; Nordmann, P. Minor extended-spectrum beta-lactamases. Clin. Microbiol. Infect. 2008, 14, 42–52, Erratum in 2008, 14, 21–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, K.; Yu, W.; Shen, P.; Lu, H.; Wang, B.; Rossen, J.W.A.; Xiao, Y. A novel Tn1696-like composite transposon (Tn6404) harboring bla IMP-4 in a Klebsiella pneumoniae isolate carrying a rare ESBL gene bla SFO-1. Sci. Rep. 2017, 7, 17321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, W.; Wachino, J.; Kimura, K.; Yamada, K.; Arakawa, Y. New plasmid-mediated aminoglycoside 6′-N-acetyltransferase, AAC(6′)-Ian, and ESBL, TLA-3, from a Serratia marcescens clinical isolate. J. Antimicrob. Chemother. 2015, 70, 1331–1337. [Google Scholar] [CrossRef] [Green Version]
- Ur Rahman, S.; Ali, T.; Ali, I.; Khan, N.A.; Han, B.; Gao, J. The Growing Genetic and Functional Diversity of Extended Spectrum Beta-Lactamases. BioMed Res. Int. 2018, 2018, 9519718. [Google Scholar] [CrossRef] [PubMed]
- Hermann, J.C.; Ridder, L.; Mulholland, A.J.; Höltje, H.D. Identification of Glu166 as the general base in the acylation reaction of class A beta-lactamases through QM/MM modeling. J. Am. Chem. Soc. 2003, 125, 9590–9591. [Google Scholar] [CrossRef]
- Meroueh, S.O.; Fisher, J.F.; Schlegel, H.B.; Mobashery, S. Ab initio QM/MM study of class A beta-lactamase acylation: Dual participation of Glu166 and Lys73 in a concerted base promotion of Ser70. J. Am. Chem. Soc. 2005, 127, 15397–15407. [Google Scholar] [CrossRef]
- Fisher, J.F.; Knowles, J.R. Bacterial Resistance to β-Lactams: The β-Lactamase. Annu. Rep. Med. Chem. 1978, 13, 239–248. [Google Scholar]
- Delmas, J.; Leyssene, D.; Dubois, D.; Birck, C.; Vazeille, E.; Robin, F.; Bonnet, R. Structural insights into substrate recognition and product expulsion in CTX-M enzymes. J. Mol. Biol. 2010, 400, 108–120. [Google Scholar] [CrossRef]
- Matagne, A.; Lamotte-Brasseur, J.; Frère, J.M. Catalytic properties of class A beta-lactamases: Efficiency and diversity. Biochem. J. 1998, 330, 581–598, Erratum in 1998, 331, 975. [Google Scholar] [CrossRef]
- Tzouvelekis, L.S.; Bonomo, R.A. SHV-type beta-lactamases. Curr. Pharm. Des. 1999, 5, 847–864. [Google Scholar]
- Huang, W.; Palzkill, T. A natural polymorphism in beta-lactamase is a global suppressor. Proc. Natl. Acad. Sci. USA 1997, 94, 8801–8806. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Minasov, G.; Shoichet, B.K. Evolution of an antibiotic resistance enzyme constrained by stability and activity trade-offs. J. Mol. Biol. 2002, 320, 85–95. [Google Scholar] [CrossRef]
- Bonnet, R. Growing group of extended-spectrum beta-lactamases: The CTX-M enzymes. Antimicrob. Agents Chemother. 2004, 48, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Andrea, M.M.; Arena, F.; Pallecchi, L.; Rossolini, G.M. CTX-M-type β-lactamases: A successful story of antibiotic resistance. Int. J. Med. Microbiol. 2013, 303, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Adamski, C.J.; Cardenas, A.M.; Brown, N.G.; Horton, L.B.; Sankaran, B.; Prasad, B.V.; Gilbert, H.F.; Palzkill, T. Molecular basis for the catalytic specificity of the CTX-M extended-spectrum β-lactamases. Biochemistry 2015, 54, 447–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marciano, D.C.; Brown, N.G.; Palzkill, T. Analysis of the plasticity of location of the Arg244 positive charge within the active site of the TEM-1 beta-lactamase. Protein Sci. 2009, 18, 2080–2089. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Delmas, J.; Sirot, J.; Shoichet, B.; Bonnet, R. Atomic resolution structures of CTX-M beta-lactamases: Extended spectrum activities from increased mobility and decreased stability. J. Mol. Biol. 2005, 348, 349–362. [Google Scholar] [CrossRef]
- Patel, M.P.; Hu, L.; Stojanoski, V.; Sankaran, B.; Prasad, B.V.V.; Palzkill, T. The Drug-Resistant Variant P167S Expands the Substrate Profile of CTX-M β-Lactamases for Oxyimino-Cephalosporin Antibiotics by Enlarging the Active Site upon Acylation. Biochemistry 2017, 56, 3443–3453. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.P.; Fryszczyn, B.G.; Palzkill, T. Characterization of the global stabilizing substitution A77V and its role in the evolution of CTX-M β-lactamases. Antimicrob. Agents Chemother. 2015, 59, 6741–6748. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.P.; Hu, L.; Brown, C.A.; Sun, Z.; Adamski, C.J.; Stojanoski, V.; Sankaran, B.; Prasad, B.V.V.; Palzkill, T. Synergistic effects of functionally distinct substitutions in β-lactamase variants shed light on the evolution of bacterial drug resistance. J. Biol. Chem. 2018, 293, 17971–17984. [Google Scholar] [CrossRef] [Green Version]
- Jacquier, H.; Marcadé, G.; Raffoux, E.; Dombret, H.; Woerther, P.L.; Donay, J.L.; Arlet, G.; Cambau, E. In vivo selection of a complex mutant TEM (CMT) from an inhibitor-resistant TEM (IRT) during ceftazidime therapy. J. Antimicrob. Chemother. 2013, 68, 2792–2796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkler, M.L.; Papp-Wallace, K.M.; Taracila, M.A.; Bonomo, R.A. Avibactam and inhibitor-resistant SHV β-lactamases. Antimicrob. Agents Chemother. 2015, 59, 3700–3709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantón, R.; Morosini, M.I.; de la Maza, O.M.; de la Pedrosa, E.G. IRT and CMT beta-lactamases and inhibitor resistance. Clin. Microbiol. Infect. 2008, 14, 53–62, Erratum in 2008, 14, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Villodres, Á.; Gil-Marqués, M.L.; Álvarez-Marín, R.; Bonnin, R.A.; Pachón-Ibáñez, M.E.; Aguilar-Guisado, M.; Naas, T.; Aznar, J.; Pachón, J.; Lepe, J.A.; et al. Extended-spectrum resistance to β-lactams/β-lactamase inhibitors (ESRI) evolved from low-level resistant Escherichia coli. J. Antimicrob. Chemother. 2020, 75, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Ding, B.; Bi, Y.; Wu, S.; Xu, S.; Xu, X.; Guo, Q.; Wang, M. CTX-M-190, a Novel β-Lactamase Resistant to Tazobactam and Sulbactam, Identified in an Escherichia coli Clinical Isolate. Antimicrob. Agents Chemother. 2016, 61, e01848-16. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Cheng, Q.; Shen, Y.; Gu, D.; Fang, Y.; Chan, E.W.; Chen, S. Genetic and Functional Characterization of blaCTX-M-199, a Novel Tazobactam and Sulbactam Resistance-Encoding Gene Located in a Conjugative mcr-1-Bearing IncI2 Plasmid. Antimicrob. Agents Chemother. 2017, 61, e00562-17. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Q.; Xu, C.; Chai, J.; Zhang, R.; Chan, E.W.; Chen, S. Structural Insight into the Mechanism of Inhibitor Resistance in CTX-M-199, a CTX-M-64 Variant Carrying the S130T Substitution. ACS Infect. Dis. 2020, 6, 577–587. [Google Scholar] [CrossRef]
- Naas, T.; Dortet, L.; Iorga, B.I. Structural and Functional Aspects of Class A Carbapenemases. Curr. Drug Targets 2016, 17, 1006–1028. [Google Scholar] [CrossRef]
- Nordmann, P.; Poirel, L. The difficult-to-control spread of carbapenemase producers among Enterobacteriaceae worldwide. Clin. Microbiol. Infect. 2014, 20, 821–830. [Google Scholar] [CrossRef] [Green Version]
- Di Luca, M.C.; Skaare, D.; Aasnaes, B.; Sundsfjord, A.; Samuelsen, Ø. Identification of a novel IMI carbapenemase variant (IMI-9) in Enterobacter cloacae complex. Int. J. Antimicrob. Agents 2016, 48, 764–765. [Google Scholar] [CrossRef]
- Poirel, L.; Ortiz de la Rosa, J.M.; Richard, A.; Aires-de-Sousa, M.; Nordmann, P. CTX-M-33, a CTX-M-15 derivative conferring reduced susceptibility to carbapenems. Antimicrob. Agents Chemother. 2019, 63, e01515-19. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.H.; Lee, J.H.; Lee, J.J.; Park, K.S.; Karim, A.M.; Lee, C.R.; Jeong, B.C.; Lee, S.H. Structural basis for carbapenem-hydrolyzing mechanisms of carbapenemases conferring antibiotic resistance. Int. J. Mol. Sci. 2015, 16, 9654–9692. [Google Scholar] [CrossRef] [PubMed]
- Queenan, A.M.; Bush, K. Carbapenemases: The versatile beta-lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majiduddin, F.K.; Palzkill, T. Amino acid residues that contribute to substrate specificity of class A beta-lactamase SME-1. Antimicrob. Agents Chemother. 2005, 49, 3421–3427. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.A.; Nossoni, Z.; Toth, M.; Stewart, N.K.; Frase, H.; Vakulenko, S.B. Role of the Conserved Disulfide Bridge in Class A Carbapenemases. J. Biol. Chem. 2016, 291, 22196–22206. [Google Scholar] [CrossRef] [Green Version]
- Mehta, S.C.; Rice, K.; Palzkill, T. Natural Variants of the KPC-2 Carbapenemase have Evolved Increased Catalytic Efficiency for Ceftazidime Hydrolysis at the Cost of Enzyme Stability. PLoS Pathog. 2015, 11, e1004949. [Google Scholar] [CrossRef]
- Mojica, M.F.; Bonomo, R.A.; Fast, W.B. Metallo-beta-Lactamases: Where do we stand? Curr. Drug Targets 2016, 17, 1029–1050. [Google Scholar] [CrossRef]
- Palzkill, T. Metallo-β-lactamase structure and function. Ann. N. Y. Acad. Sci. 2013, 1277, 91–104. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. Interplay between β-lactamases and new β-lactamase inhibitors. Nat. Rev. Microbiol. 2019, 17, 295–306, Erratum in 2019, 17, 459–460. [Google Scholar] [CrossRef]
- Wu, W.; Feng, Y.; Tang, G.; Qiao, F.; McNally, A.; Zong, Z. NDM Metallo-β-Lactamases and Their Bacterial Producers in Health Care Settings. Clin. Microbiol. Rev. 2019, 32, e00115-18. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.H.; Hu, Z.Q. Epidemiology and genetics of VIM-type metallo-β-lactamases in Gram-negative bacilli. Future Microbiol. 2011, 6, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.H.; Hu, Z.Q. IMP-type metallo-β-lactamases in Gram-negative bacilli: Distribution, phylogeny, and association with integrons. Crit. Rev. Microbiol. 2011, 37, 214–226. [Google Scholar] [CrossRef]
- Feng, H.; Liu, X.; Wang, S.; Fleming, J.; Wang, D.C.; Liu, W. The mechanism of NDM-1-catalyzed carbapenem hydrolysis is distinct from that of penicillin or cephalosporin hydrolysis. Nat. Commun. 2017, 8, 2242. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Hu, L.; Sankaran, B.; Prasad, B.V.V.; Palzkill, T. Differential active site requirements for NDM-1 β-lactamase hydrolysis of carbapenem versus penicillin and cephalosporin antibiotics. Nat. Commun. 2018, 9, 4524. [Google Scholar] [CrossRef]
- Palacios, A.R.; Mojica, M.F.; Giannini, E.; Taracila, M.A.; Bethel, C.R.; Alzari, P.M.; Otero, L.H.; Klinke, S.; Llarrull, L.I.; Bonomo, R.A.; et al. The Reaction Mechanism of Metallo-β-Lactamases Is Tuned by the Conformation of an Active-Site Mobile Loop. Antimicrob. Agents Chemother. 2018, 63, e01754-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Docquier, J.D.; Lamotte-Brasseur, J.; Galleni, M.; Amicosante, G.; Frère, J.M.; Rossolini, G.M. On functional and structural heterogeneity of VIM-type metallo-beta-lactamases. J. Antimicrob. Chemother. 2003, 51, 257–266. [Google Scholar] [CrossRef]
- Borra, P.S.; Leiros, H.K.; Ahmad, R.; Spencer, J.; Leiros, I.; Walsh, T.R.; Sundsfjord, A.; Samuelsen, O. Structural and computational investigations of VIM-7: Insights into the substrate specificity of vim metallo-β-lactamases. J. Mol. Biol. 2011, 411, 174–189. [Google Scholar] [CrossRef]
- Leiros, H.K.; Edvardsen, K.S.; Bjerga, G.E.; Samuelsen, Ø. Structural and biochemical characterization of VIM-26 shows that Leu224 has implications for the substrate specificity of VIM metallo-β-lactamases. FEBS J. 2015, 282, 1031–1042. [Google Scholar] [CrossRef] [Green Version]
- Salimraj, R.; Hinchliffe, P.; Kosmopoulou, M.; Tyrrell, J.M.; Brem, J.; van Berkel, S.S.; Verma, A.; Owens, R.J.; McDonough, M.A.; Walsh, T.R.; et al. Crystal structures of VIM-1 complexes explain active site heterogeneity in VIM-class metallo-β-lactamases. FEBS J. 2019, 286, 169–183. [Google Scholar] [CrossRef]
- Leonard, D.A.; Bonomo, R.A.; Powers, R.A. Class D β-lactamases: A reappraisal after five decades. Acc. Chem. Res. 2013, 46, 2407–2415. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Potron, A.; Nordmann, P. OXA-48-like carbapenemases: The phantom menace. J. Antimicrob. Chemother. 2012, 67, 1597–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitout, J.D.D.; Peirano, G.; Kock, M.M.; Strydom, K.A.; Matsumura, Y. The Global Ascendency of OXA-48-Type Carbapenemases. Clin. Microbiol. Rev. 2019, 33, e00102-19. [Google Scholar] [CrossRef] [PubMed]
- Schneider, K.D.; Bethel, C.R.; Distler, A.M.; Hujer, A.M.; Bonomo, R.A.; Leonard, D.A. Mutation of the active site carboxy-lysine (K70) of OXA-1 beta-lactamase results in a deacylation-deficient enzyme. Biochemistry 2009, 48, 6136–6145. [Google Scholar] [CrossRef] [Green Version]
- Verma, V.; Testero, S.A.; Amini, K.; Wei, W.; Liu, J.; Balachandran, N.; Monoharan, T.; Stynes, S.; Kotra, L.P.; Golemi-Kotra, D. Hydrolytic mechanism of OXA-58 enzyme, a carbapenem-hydrolyzing class D β-lactamase from Acinetobacter baumannii. J. Biol. Chem. 2011, 286, 37292–37303. [Google Scholar] [CrossRef] [Green Version]
- Santillana, E.; Beceiro, A.; Bou, G.; Romero, A. Crystal structure of the carbapenemase OXA-24 reveals insights into the mechanism of carbapenem hydrolysis. Proc. Natl. Acad. Sci. USA 2007, 104, 5354–5359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.A.; Antunes, N.T.; Stewart, N.K.; Toth, M.; Kumarasiri, M.; Chang, M.; Mobashery, S.; Vakulenko, S.B. Structural basis for carbapenemase activity of the OXA-23 β-lactamase from Acinetobacter baumannii. Chem. Biol. 2013, 20, 1107–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Docquier, J.D.; Calderone, V.; De Luca, F.; Benvenuti, M.; Giuliani, F.; Bellucci, L.; Tafi, A.; Nordmann, P.; Botta, M.; Rossolini, G.M.; et al. Crystal structure of the OXA-48 beta-lactamase reveals mechanistic diversity among class D carbapenemases. Chem. Biol. 2009, 16, 540–547. [Google Scholar] [CrossRef]
- Smith, C.A.; Antunes, N.T.; Toth, M.; Vakulenko, S.B. Crystal structure of carbapenemase OXA-58 from Acinetobacter baumannii. Antimicrob. Agents Chemother. 2014, 58, 2135–2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.A.; Stewart, N.K.; Toth, M.; Vakulenko, S.B. Structural Insights into the Mechanism of Carbapenemase Activity of the OXA-48 β-Lactamase. Antimicrob. Agents Chemother. 2019, 63, e01202-19. [Google Scholar] [CrossRef] [Green Version]
- Drawz, S.M.; Bonomo, R.A. Three decades of beta-lactamase inhibitors. Clin. Microbiol. Rev. 2010, 23, 160–201. [Google Scholar] [CrossRef] [Green Version]
- Nordmann, P.; Mammeri, H. Extended-spectrum cephalosporinases: Structure, detection and epidemiology. Future Microbiol. 2007, 2, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Bauernfeind, A.; Chong, Y.; Schweighart, S. Extended broad spectrum beta-lactamase in Klebsiella pneumoniae including resistance to cephamycins. Infection 1989, 17, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Tamma, P.D.; Doi, Y.; Bonomo, R.A.; Johnson, J.K.; Simner, P.J.; Antibacterial Resistance Leadership Group. A Primer on AmpC β-Lactamases: Necessary Knowledge for an Increasingly Multidrug-resistant World. Clin. Infect. Dis. 2019, 69, 1446–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pires, J.; Taracila, M.; Bethel, C.R.; Doi, Y.; Kasraian, S.; Tinguely, R.; Sendi, P.; Bonomo, R.A.; Endimiani, A. In Vivo Evolution of CMY-2 to CMY-33 β-Lactamase in Escherichia coli Sequence Type 131: Characterization of an Acquired Extended-Spectrum AmpC Conferring Resistance to Cefepime. Antimicrob. Agents Chemother. 2015, 59, 7483–7488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donà, V.; Scheidegger, M.; Pires, J.; Furrer, H.; Atkinson, A.; Babouee Flury, B. Gradual in vitro Evolution of Cefepime Resistance in an ST131 Escherichia coli Strain Expressing a Plasmid-Encoded CMY-2 β-Lactamase. Front. Microbiol. 2019, 10, 1311. [Google Scholar] [CrossRef] [Green Version]
- Castanheira, M.; Farrell, S.E.; Deshpande, L.M.; Mendes, R.E.; Jones, R.N. Prevalence of β-lactamase-encoding genes among Enterobacteriaceae bacteremia isolates collected in 26 U.S. hospitals: Report from the SENTRY Antimicrobial Surveillance Program (2010). Antimicrob. Agents Chemother. 2013, 57, 3012–3020. [Google Scholar] [CrossRef] [Green Version]
- Peirano, G.; Pitout, J.D.D. Extended-Spectrum β-Lactamase-Producing Enterobacteriaceae: Update on Molecular Epidemiology and Treatment Options. Drugs 2019, 79, 1529–1541. [Google Scholar] [CrossRef]
- Public Health England. English Surveillance Programme for Antimicrobial Utilisation and Resistance (ESPAUR) Report 2017; Public Health England: London, UK, 2017. Available online: https://www.gov.uk/government/publications/english-surveillance-programme-antimicrobial-utilisation-and-resistance-espaur-report (accessed on 26 June 2020).
- Robin, F.; Beyrouthy, R.; Bonacorsi, S.; Aissa, N.; Bret, L.; Brieu, N.; Cattoir, V.; Chapuis, A.; Chardon, H.; Degand, N.; et al. Inventory of extended-spectrum-b-lactamase-producing Enterobacteriaceae in France as assessed by a multicenter study. Antimicrob. Agents Chemother. 2017, 61, e01911-16. [Google Scholar] [CrossRef] [Green Version]
- Rios, E.; Lopez, M.C.; Rodriguez-Avial, I.; Culebras, E.; Picazo, J.J. Detection of Escherichia coli ST131 clonal complex (ST705) and Klebsiella pneumoniae ST15 among faecal carriage of extended spectrum b-lactamase- and carbapenemase-producing Enterobacteriaceae. J. Med. Microbiol. 2017, 66, 169–174. [Google Scholar] [CrossRef]
- Pietsch, M.; Eller, C.; Wendt, C.; Holfelder, M.; Falgenhauer, L.; Fruth, A.; Grössl, T.; Leistner, R.; Valenza, G.; Werner, G.; et al. Molecular characterisation of extended-spectrum b-lactamase (ESBL)-producing Escherichia coli isolates from hospital and ambulatory patients in Germany. Vet. Microbiol. 2017, 200, 130–137. [Google Scholar] [CrossRef]
- Lee, M.Y.; Ko, K.S.; Kang, C.I.; Chung, D.R.; Peck, K.R.; Song, J.H. High prevalence of CTX-M-15-producing Klebsiella pneumoniae isolates in Asian countries: Diverse clones and clonal dissemination. Int. J. Antimicrob. Agents 2011, 38, 160–163. [Google Scholar] [CrossRef] [PubMed]
- Quan, J.; Zhao, D.; Liu, L.; Chen, Y.; Zhou, J.; Akova, M.; Yu, Y. High prevalence of ESBL-producing Escherichia coli and Klebsiella pneumoniae in community-onset bloodstream infections in China. J. Antimicrob. Chemother. 2017, 72, 273–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musicha, P.; Feasey, N.A.; Cain, A.K.; Kallonen, T.; Chaguza, C.; Peno, C.; Khonga, M.; Thompson, S.; Gray, K.J.; Mather, A.E.; et al. Genomic landscape of extended-spectrum b-lactamase resistance in Escherichia coli from an urban African setting. J. Antimicrob. Chemother. 2017, 72, 1602–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Johnston, B.; Clabots, C.; Kuskowski, M.A.; Castanheira, M. Escherichia coli sequence type ST131 as the major cause of serious multidrug-resistant E. coli infections in the United States. Clin. Infect. Dis. 2010, 51, 286–294. [Google Scholar] [CrossRef] [Green Version]
- Doi, Y.; Park, Y.S.; Rivera, J.I.; Adams-Haduch, J.M.; Hingwe, A.; Sordillo, E.M.; Lewis, J.S.; Howard, W.J.; Johnson, L.E.; Polsky, B.; et al. Community-associated extended-spectrum β-lactamase producing Escherichia coli infection in the United States. Clin. Infect. Dis. 2013, 56, 641–648. [Google Scholar] [CrossRef]
- Munoz-Price, L.S.; Poirel, L.; Bonomo, R.A.; Schwaber, M.J.; Daikos, G.L.; Cormican, M.; Cornaglia, G.; Garau, J.; Gniadkowski, M.; Hayden, M.K.; et al. Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet Infect. Dis. 2013, 13, 785–796. [Google Scholar] [CrossRef] [Green Version]
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob. Agents Chemother. 2009, 53, 5046. [Google Scholar] [CrossRef] [Green Version]
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Doumith, M.; Giske, C.G.; Irfan, S.; et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: A molecular, biological, and epidemiological study. Lancet Infect. Dis. 2010, 10, 597. [Google Scholar] [CrossRef]
- Woodworth, K.R.; Walters, M.S.; Weiner, L.M.; Edwards, J.; Brown, A.C.; Huang, J.Y.; Malik, S.; Slayton, R.B.; Paul, P.; Capers, C.; et al. Vital Signs: Containment of Novel Multidrug-Resistant Organisms and Resistance Mechanisms—United States, 2006–2017. MMWR 2018, 67, 396. [Google Scholar] [CrossRef] [Green Version]
- Dortet, L.; Poirel, L.; Nordmann, P. Worldwide dissemination of the NDM-type carbapenemases in Gram-negative bacteria. BioMed Res. Int. 2014, 2014, 249856. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.; Fu, Y.; Zhang, Q.; Qi, H.; Wen, J.G.; Xu, H.; Xu, L.; Zeng, L.; Tian, H.; Rong, L.; et al. High incidence and endemic spread of NDM-1-positive Enterobacteriaceae in Henan Province, China. Antimicrob. Agents Chemother. 2014, 58, 4275–4282. [Google Scholar] [CrossRef] [Green Version]
- Tavoschi, L.; Forni, S.; Porretta, A.; Righi, L.; Pieralli, F.; Menichetti, F.; Falcone, M.; Gemignani, G.; Sani, S.; Vivani, P.; et al. Prolonged outbreak of New Delhi metallo-beta-lactamase-producing carbapenem-resistant Enterobacterales (NDM-CRE), Tuscany, Italy, 2018 to 2019. Euro Surveill. 2020, 25, 2000085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albiger, B.; Glasner, C.; Struelens, M.J.; Grundmann, H.; Monnet, D.L.; European Survey of Carbapenemase-Producing Enterobacteriaceae (EuSCAPE) working group. Carbapenemase-producing Enterobacteriaceae in Europe: Assessment by national experts from 38 countries, May 2015. Euro Surveill. 2015, 20, 30062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, G.; Bonomo, R.A. “Stormy waters ahead”: Global emergence of carbapenemases. Front. Microbiol. 2013, 4, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, T.R.; Toleman, M.A.; Poirel, L.; Nordmann, P. Metallo-ß-Lactamases: The Quiet before the Storm? Clin. Microbiol. Rev. 2005, 18, 306–325. [Google Scholar] [CrossRef] [Green Version]
- Logan, L.K.; Weinstein, R.A. The Epidemiology of Carbapenem-Resistant Enterobacteriaceae: The Impact and Evolution of a Global Menace. J. Infect. Dis. 2017, 215, S28–S36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumura, Y.; Peirano, G.; Devinney, R.; Bradford, P.A.; Motyl, M.R.; Adams, M.D.; Chen, L.; Kreiswirth, B.; Pitout, J.D.D. Genomic epidemiology of global VIM-producing Enterobacteriaceae. J. Antimicrob. Chemother. 2017, 72, 2249–2258. [Google Scholar] [CrossRef] [Green Version]
- Sidjabat, H.E.; Townell, N.; Nimmo, G.R.; George, N.M.; Robson, J.; Vohra, R.; Davis, L.; Heney, C.; Paterson, D.L. Dominance of IMP-4-producing Enterobacter cloacae among carbapenemase- producing Enterobacteriaceae in Australia. Antimicrob. Agents Chemother. 2015, 59, 4059–4066. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, Y.; Funaki, T.; Yasuhara, T.; Sugano, E.; Ugajin, K.; Tahara, S.; Fukuchi, K. Molecular characteristics of a carbapenemase-producing Enterobacter species and Klebsiella species outbreak in a Japanese University Hospital. Showa Univ. J. Med. Sci. 2017, 29, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Nordmann, P.; Naas, T.; Poirel, L. Global spread of carbapenemase-producing Enterobacteriaceae. Emerg. Infect. Dis. 2011, 17, 1791–1798. [Google Scholar] [CrossRef]
- Iovleva, A.; Doi, Y. Carbapenem-Resistant Enterobacteriaceae. Clin. Lab. Med. 2017, 37, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, L.; Dortet, L.; Cotellon, G.; Creton, E.; Cuzon, G.; Ponties, V.; Bonnin, R.A.; Naas, T. Diversity of carbapenemase-producing Escherichia coli isolates in France in 2012–2013. Antimicrob. Agents Chemother. 2018, 62, e00266-18. [Google Scholar] [CrossRef] [Green Version]
- Ortega, A.; Saez, D.; Bautista, V.; Fernandez-Romero, S.; Lara, N.; Aracil, B.; Perez-Vazquez, M.; Campos, J.; Oteo, J.; Spanish Collaborating Group for the Antibiotic Resistance Surveillance Programme. Carbapenemase-producing Escherichia coli is becoming more prevalent in Spain mainly because of the polyclonal dissemination of OXA-48. J. Antimicrob. Chemother. 2016, 71, 2131–2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balkan, I.I.; Aygün, G.; Aydın, S.; Mutcalı, S.I.; Kara, Z.; Kuşkucu, M.; Midilli, K.; Şemen, V.; Aras, S.; Yemişen, M.; et al. Blood stream infections due to OXA-48-like carbapenemase-producing Enterobacteriaceae: Treatment and survival. Int. J. Infect. Dis. 2014, 26, 51–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castanheira, M.; Deshpande, L.M.; Mathai, D.; Bell, J.M.; Jones, R.N.; Mendes, R.E. Early dissemination of NDM-1- and OXA-181-producing Enterobacteriaceae in Indian hospitals: Report from the SENTRY antimicrobial surveillance program, 2006–2007. Antimicrob. Agents Chemother. 2011, 55, 1274–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamal, D.; Fernandez-Martínez, M.; El-Defrawy, I.; Ocampo-Sosa, A.A.; Martínez-Martínez, L. First identification of NDM-5 associated with OXA-181 in Escherichia coli from Egypt. Emerg. Microbes Infect. 2016, 5, e30. [Google Scholar] [CrossRef] [Green Version]
- Overballe-Petersen, S.; Roer, L.; Ng, K.; Hansen, F.; Justesen, U.S.; Andersen, L.P.; Stegger, M.; Hammerum, A.M.; Hasman, H. Complete nucleotide sequence of an Escherichia coli sequence type 410 strain carrying blaNDM-5 on an IncF multidrug resistance plasmid and blaOXA-181 on an IncX3 Plasmid. Genome. Announc. 2018, 6, e01542-17. [Google Scholar] [CrossRef] [Green Version]
- Jacoby, G.A. AmpC beta-lactamases. Clin. Microbiol. Rev. 2009, 22, 161–182. [Google Scholar] [CrossRef] [Green Version]
- Doi, Y.; Paterson, D.L.; Adams-Haduch, J.M.; Sidjabat, H.E.; O’Keefe, A.; Endimiani, A.; Bonomo, R.A. Reduced susceptibility to cefepime among Escherichia coli clinical isolates producing novel variants of CMY-2 beta-lactamase. Antimicrob. Agents Chemother. 2009, 53, 3159–3161. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, B.; Mukherjee, M. Emergence of co-production of plasmid-mediated AmpC beta-lactamase and ESBL in cefoxitin-resistant uropathogenic Escherichia coli. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1449–1454. [Google Scholar] [CrossRef]
- Wellington, E.M.H.; Boxall, A.B.; Cross, P.; Feil, E.J.; Gaze, W.H.; Hawkey, P.M.; Johnson-Rollings, A.S.; Jones, D.L.; Lee, N.M.; Otten, W.; et al. The role of the natural environment in the emergence of antibiotic resistance in gramnegative bacteria. Lancet Infect. Dis. 2013, 13, 155–165. [Google Scholar] [CrossRef]
- Gomi, R.; Matsuda, T.; Matsumura, Y.; Yamamoto, M.; Tanaka, M.; Ichiyama, S.; Yoneda, M. Occurrence of Clinically Important Lineages, Including the Sequence Type 131 C1-M27 Subclone, among Extended-Spectrum-β-Lactamase-Producing Escherichia coli in Wastewater. Antimicrob. Agents Chemother. 2017, 61, e00564-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotay, S.; Chai, W.; Guilford, W.; Barry, K.; Mathers, A.J. Spread from the Sink to the Patient: In Situ Study Using Green Fluorescent Protein (GFP)-Expressing Escherichia coli To Model Bacterial Dispersion from Hand-Washing Sink-Trap Reservoirs. Appl. Environ. Microbiol. 2017, 83, e03327-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammerum, A.M.; Larsen, J.; Andersen, V.D.; Lester, C.H.; Skovgaard Skytte, T.S.; Hansen, F.; Olsen, S.S.; Mordhorst, H.; Skov, R.L.; Aarestrup, F.M.; et al. Characterization of extended-spectrumβ-lactamase (ESBL)-producing Escherichia coli obtained from Danish pigs, pig farmers and their families from farms with high or no consumption of third- or fourth-generation cephalosporins. J. Antimicrob. Chemother. 2014, 69, 2650–2657. [Google Scholar] [CrossRef] [PubMed]
- Liebana, E.; Carattoli, A.; Coque, T.M.; Hasman, H.; Magiorakos, A.P.; Mevius, D.; Peixe, L.; Poirel, L.; Schuepbach-Regula, G.; Torneke, K.; et al. Public health risks of enterobacterial isolates producing extended-spectrumβ-lactamases or AmpCβ-lactamases in food and food-producing animals: An EU perspective of epidemiology, analytical methods, risk factors, and control options. Clin. Infect. Dis. 2013, 56, 1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pehrsson, E.C.; Tsukayama, P.; Patel, S.; Mejía-Bautista, M.; Sosa-Soto, G.; Navarrete, K.M.; Calderon, M.; Cabrera, L.; Hoyos-Arango, W.; Bertoli, M.T.; et al. Interconnected microbiomes and resistomes in low-income human habitats. Nature 2016, 533, 212–216. [Google Scholar] [CrossRef] [Green Version]
- Chong, Y.; Shimoda, S.; Shimono, N. Current epidemiology, genetic evolution and clinical impact of extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella pneumoniae. Infect. Genet. Evol. 2018, 61, 185–188. [Google Scholar] [CrossRef]
- Kelly, A.M.; Mathema, B.; Larson, E.L. Carbapenem-resistant Enterobacteriaceae in the community: A scoping review. Int. J. Antimicrob. Agents 2017, 50, 127–134. [Google Scholar] [CrossRef]
- Szmolka, A.; Nagy, B. Multidrug resistant commensal Escherichia coli in animals and its impact for public health. Front. Microbiol. 2013, 4, 258. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.U.; Maryam, L.; Zarrilli, R. Structure, Genetics and Worldwide Spread of New Delhi Metallo-β-lactamase (NDM): A threat to public health. BMC Microbiol. 2017, 17, 101. [Google Scholar] [CrossRef] [Green Version]
- Bakthavatchalam, Y.D.; Anandan, S.; Veeraraghavan, B. Laboratory Detection and Clinical Implication of Oxacillinase-48 like Carbapenemase: The Hidden Threat. J. Glob. Infect. Dis. 2016, 8, 41–50. [Google Scholar] [PubMed]
- Park, S.H.; Byun, J.H.; Choi, S.M.; Lee, D.G.; Kwon, J.; Kim, S.H.; Kwon, J.C.; Park, C.; Choi, J.H.; Yoo, J.H. Molecular epidemiology of extended-spectrum β-lactamase-producing Escherichia coli in the community and hospital in Korea: Emergence of ST131 producing CTX-M-15. BMC Infect. Dis. 2012, 12, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumura, Y.; Pitout, J.D.D.; Peirano, G.; DeVinney, R.; Noguchi, T.; Yamamoto, M.; Gomi, R.; Matsuda, T.; Nakano, S.; Nagao, M.; et al. Rapid identification of different Escherichia coli sequence type 131 clades. Antimicrob. Agents Chemother. 2017, 61, e00179-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, D.D.; Wan, L.G.; Deng, Q.; Liu, Y. Emergence of KPC-producing Klebsiella pneumoniae hypervirulent clone of capsular serotype K1 that belongs to sequence type 11 in Mainland China. Diagn. Microbiol. Infect. Dis. 2016, 85, 192–194. [Google Scholar] [CrossRef] [PubMed]
- Kazi, M.; Drego, L.; Nikam, C.; Ajbani, K.; Soman, R.; Shetty, A.; Rodrigues, C. Molecular characterization of carbapenem-resistant Enterobacteriaceae at a tertiary care laboratory in Mumbai. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, P.; Meenakshisundaram, J.; Jayaraman, P. blaKPC gene Detection in Clinical Isolates of Carbapenem Resistant Enterobacteriaceae in a Tertiary Care Hospital. J. Clin. Diagn. Res. 2013, 7, 2736–2738. [Google Scholar]
- Van Duin, D.; Doi, Y. The global epidemiology of carbapenemase-producing Enterobacteriaceae. Virulence 2017, 8, 460–469. [Google Scholar] [CrossRef]
- Mushi, M.F.; Mshana, S.E.; Imirzalioglu, C.; Bwanga, F. Carbapenemase genes among multidrug resistant gram negative clinical isolates from a tertiary hospital in Mwanza, Tanzania. BioMed Res. Int. 2014, 2014, 303104. [Google Scholar] [CrossRef] [Green Version]
- Brink, A.J.; Coetzee, J.; Clay, C.G.; Sithole, S.; Richards, G.A.; Poirel, L.; Nordmann, P. Emergence of New Delhi metallo-beta-lactamase (NDM-1) and Klebsiella pneumoniae carbapenemase (KPC-2) in South Africa. J. Clin. Microbiol. 2012, 50, 525–527. [Google Scholar] [CrossRef] [Green Version]
- Pitout, J.D.; Nordmann, P.; Poirel, L. Carbapenemase-producing Klebsiella pneumoniae, a key pathogen set for global nosocomial dominance. Antimicrob. Agents Chemother. 2015, 59, 5873–5884. [Google Scholar] [CrossRef] [Green Version]
- Ho, P.L.; Cheung, Y.Y.; Lo, W.U.; Li, Z.; Chow, K.H.; Lin, C.H.; Chan, J.F.; Cheng, V.C. Molecular characterization of an atypical IncX3 plasmid pKPC-NY79 carrying blaKPC-2 in a Klebsiella pneumoniae. Curr. Microbiol. 2013, 67, 493–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekizuka, T.; Matsui, M.; Yamane, K.; Takeuchi, F.; Ohnishi, M.; Hishinuma, A.; Arakawa, Y.; Kuroda, M. Complete sequencing of the blaNDM-1-positive IncA/C plasmid from Escherichia coli ST38 isolate suggests a possible origin from plant pathogens. PLoS ONE 2011, 6, e25334. [Google Scholar] [CrossRef]
- Villa, L.; Poirel, L.; Nordmann, P.; Carta, C.; Carattoli, A. Complete sequencing of an IncH plasmid carrying the blaNDM-1, blaCTX-M-15 and qnrB1 genes. J. Antimicrob. Chemother. 2012, 67, 1645–1650. [Google Scholar] [CrossRef]
- Wailan, A.M.; Sartor, A.L.; Zowawi, H.M.; Perry, J.D.; Paterson, D.L.; Sidjabat, H.E. Genetic contexts of blaNDM-1 in patients carrying multiple NDM-producing strains. Antimicrob. Agents Chemother. 2015, 59, 7405–7410. [Google Scholar] [CrossRef] [Green Version]
- Giske, C.G.; Fröding, I.; Hasan, C.M.; Turlej-Rogacka, A.; Toleman, M.; Livermore, D.; Woodford, N.; Walsh, T.R. Diverse sequence types of Klebsiella pneumoniae contribute to the dissemination of blaNDM-1 in India, Sweden, and the United Kingdom. Antimicrob. Agents Chemother. 2012, 56, 2735–2738. [Google Scholar] [CrossRef] [Green Version]
- Khong, W.X.; Xia, E.; Marimuthu, K.; Xu, W.; Teo, Y.Y.; Tan, E.L.; Neo, S.; Krishnan, P.U.; Ang, B.S.; Lye, D.C.; et al. Local transmission and global dissemination of New Delhi metallo-b-lactamase (NDM): A whole genome analysis. BMC Genom. 2016, 17, 452. [Google Scholar] [CrossRef] [Green Version]
- Wailan, A.M.; Sidjabat, H.E.; Yam, W.K.; Alikhan, N.F.; Petty, N.K.; Sartor, A.L.; Williamson, D.A.; Forde, B.M.; Schembri, M.A.; Beatson, S.A.; et al. Mechanisms involved in acquisition of blaNDM genes by IncA/C2 and IncFIIY plasmids. Antimicrob. Agents Chemother. 2016, 60, 4082–4088. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A. Plasmids and the spread of resistance. Int. J. Med. Microbiol. 2013, 303, 298–304. [Google Scholar] [CrossRef]
- Espedido, B.A.; Partridge, S.R.; Iredell, J.R. bla(IMP-4) in different genetic contexts in Enterobacteriaceae isolates from Australia. Antimicrob. Agents Chemother. 2008, 52, 2984–2987. [Google Scholar] [CrossRef] [Green Version]
- Walsh, T.R. Clinically significant carbapenemases: An update. Curr. Opin. Infect. Dis. 2008, 21, 367–371. [Google Scholar] [CrossRef]
- Giani, T.; Conte, V.; di Pilato, V.; Aschbacher, R.; Weber, C.; Weber, C.; Larcher, C.; Gian Maria Rossolini, G.M. Escherichia coli from Italy producing OXA-48 carbapenemase encoded by a novel Tn1999 transposon derivative. Antimicrob. Agents Chemother. 2012, 56, 2211–2213. [Google Scholar] [CrossRef] [Green Version]
- Potron, A.; Poirel, L.; Rondinaud, E.; Nordmann, P. Intercontinental spread of OXA-48 b-lactamase-producing Enterobacteriaceae over a 11-year period, 2001 to 2011. Euro Surveill. 2013, 18, 20549. [Google Scholar] [CrossRef]
- Solgi, H.; Giske, C.G.; Badmasti, F.; Aghamohammad, S.; Havaei, S.A.; Sabeti, S.; Mostafavizadeh, K.; Shahcheraghi, F. Emergence of carbapenem resistant Escherichia coli isolates producing blaNDM and blaOXA-48-like carried on IncA/C and IncL/M plasmids at two Iranian university hospitals. Infect. Genet. Evol. 2017, 55, 318–323. [Google Scholar] [CrossRef]
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M.; et al. Attributable deaths and disability-adjusted lifeyears caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: A population-level modelling analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Tumbarello, M.; Sanguinetti, M.; Montuori, E.; Trecarichi, E.M.; Posteraro, B.; Fiori, B.; Citton, R.; D’Inzeo, T.; Fadda, G.; Cauda, R.; et al. Predictors of mortality in patients with bloodstream infections caused by extended-spectrum-beta-lactamase-producing Enterobacteriaceae: Importance of inadequate initial antimicrobial treatment. Antimicrob. Agents Chemother. 2007, 51, 1987–1994. [Google Scholar] [CrossRef] [Green Version]
- Tumbarello, M.; Sali, M.; Trecarichi, E.M.; Leone, F.; Rossi, M.; Fiori, B.; De Pascale, G.; D’Inzeo, T.; Sanguinetti, M.; Fadda, G.; et al. Bloodstream infections caused by extended-spectrum-beta-lactamase- producing Escherichia coli: Risk factors for inadequate initial antimicrobial therapy. Antimicrob. Agents Chemother. 2008, 52, 3244–3252. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Gutiérrez, B.; Salamanca, E.; de Cueto, M.; Hsueh, P.R.; Viale, P.; Paño-Pardo, J.R.; Venditti, M.; Tumbarello, M.; Daikos, G.; Pintado, V.; et al. A Predictive Model of Mortality in Patients With Bloodstream Infections due to Carbapenemase-Producing Enterobacteriaceae. Mayo Clin. Proc. 2016, 91, 1362–1371. [Google Scholar] [CrossRef] [Green Version]
- Palacios-Baena, Z.R.; Gutiérrez-Gutiérrez, B.; De Cueto, M.; Viale, P.; Venditti, M.; Hernández-Torres, A.; Oliver, A.; Martínez-Martínez, L.; Calbo, E.; Pintado, V.; et al. Development and validation of the INCREMENT-ESBL predictive score for mortality in patients with bloodstream infections due to extended-spectrum-β-lactamase-producing Enterobacteriaceae. J. Antimicrob. Chemother. 2017, 72, 906–913. [Google Scholar] [CrossRef] [Green Version]
- Tumbarello, M.; Spanu, T.; Di Bidino, R.; Marchetti, M.; Ruggeri, M.; Trecarichi, E.M.; De Pascale, G.; Proli, E.M.; Cauda, R.; Cicchetti, A.; et al. Costs of bloodstream infections caused by Escherichia coli and influence of extended-spectrum-beta-lactamase production and inadequate initial antibiotic therapy. Antimicrob. Agents Chemother. 2010, 54, 4085–4091. [Google Scholar] [CrossRef] [Green Version]
- Shamsrizi, P.; Gladstone, B.P.; Carrara, E.; Luise, D.; Cona, A.; Bovo, C.; Tacconelli, E. Variation of effect estimates in the analysis of mortality and length of hospital stay in patients with infections caused by bacteria-producing extended-spectrum beta-lactamases: A systematic review and meta-analysis. BMJ Open 2020, 10, e030266. [Google Scholar] [CrossRef] [Green Version]
- Harris, P.N.A.; Tambyah, P.A.; Lye, D.C.; Mo, Y.; Lee, T.H.; Yilmaz, M.; Alenazi, T.H.; Arabi, Y.; Falcone, M.; Bassetti, M.; et al. Effect of Piperacillin-Tazobactam vs Meropenem on 30-Day Mortality for Patients with E coli or Klebsiella pneumoniae Bloodstream Infection and Ceftriaxone Resistance: A Randomized Clinical Trial. JAMA 2018, 320, 984–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhammed, M.; Flokas, M.E.; Detsis, M.; Alevizakos, M.; Mylonakis, E. Comparison Between Carbapenems and β-Lactam/β-Lactamase Inhibitors in the Treatment for Bloodstream Infections Caused by Extended-Spectrum β-Lactamase-Producing Enterobacteriaceae: A Systematic Review and Meta-Analysis. Open Forum Infect. Dis. 2017, 4, ofx099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassetti, M.; Vena, A.; Giacobbe, D.R.; Falcone, M.; Tiseo, G.; Giannella, M.; Pascale, R.; Meschiari, M.; Digaetano, M.; Oliva, A.; et al. Ceftolozane/Tazobactam for Treatment of Severe ESBL-Producing Enterobacterales Infections: A Multicenter Nationwide Clinical Experience (CEFTABUSE II Study). Open Forum Infect. Dis. 2020, 7, ofaa139. [Google Scholar] [CrossRef] [PubMed]
- Vena, A.; Giacobbe, D.R.; Castaldo, N.; Cattelan, A.; Mussini, C.; Luzzati, R.; Rosa, F.G.; Del Puente, F.; Mastroianni, C.M.; Cascio, A.; et al. Clinical Experience with Ceftazidime-Avibactam for the Treatment of Infections due to Multidrug-Resistant Gram-Negative Bacteria Other than Carbapenem-Resistant Enterobacterales. Antibiotics 2020, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Tumbarello, M.; Viale, P.; Viscoli, C.; Trecarichi, E.M.; Tumietto, F.; Marchese, A.; Spanu, T.; Ambretti, S.; Ginocchio, F.; Cristini, F.; et al. Predictors of mortality in bloodstream infections caused by Klebsiella pneumoniae carbapenemase-producing K. pneumoniae: Importance of combination therapy. Clin. Infect. Dis. 2012, 55, 943–950. [Google Scholar] [CrossRef] [Green Version]
- Shields, R.K.; Nguyen, M.H.; Chen, L.; Press, E.G.; Potoski, B.A.; Marini, R.V.; Doi, Y.; Kreiswirth, B.N.; Clancy, C.J. Ceftazidime-Avibactam Is Superior to Other Treatment Regimens against Carbapenem-Resistant Klebsiella pneumoniae Bacteremia. Antimicrob. Agents Chemother. 2017, 61, e00883-17. [Google Scholar] [CrossRef] [Green Version]
- Tumbarello, M.; Trecarichi, E.M.; Corona, A.; De Rosa, F.G.; Bassetti, M.; Mussini, C.; Menichetti, F.; Viscoli, C.; Campoli, C.; Venditti, M.; et al. Efficacy of Ceftazidime-Avibactam Salvage Therapy in Patients With Infections Caused by Klebsiella pneumoniae Carbapenemase-producing K. pneumoniae. Clin. Infect. Dis. 2019, 68, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Martin, A.; Fahrbach, K.; Zhao, Q.; Lodise, T. Association Between Carbapenem Resistance and Mortality Among Adult, Hospitalized Patients With Serious Infections Due to Enterobacteriaceae: Results of a Systematic Literature Review and Meta-analysis. Open Forum Infect. Dis. 2018, 5, ofy150. [Google Scholar] [CrossRef]
- Tumbarello, M.; Trecarichi, E.M.; De Rosa, F.G.; Giannella, M.; Giacobbe, D.R.; Bassetti, M.; Losito, A.R.; Bartoletti, M.; Del Bono, V.; Corcione, S.; et al. Infections caused by KPC-producing Klebsiella pneumoniae: Differences in therapy and mortality in a multicentre study. J. Antimicrob. Chemother. 2015, 70, 2133–2143. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Gutiérrez, B.; Salamanca, E.; de Cueto, M.; Hsueh, P.R.; Viale, P.; Paño-Pardo, J.R.; Venditti, M.; Tumbarello, M.; Daikos, G.; Cantón, R.; et al. Effect of appropriate combination therapy on mortality of patients with bloodstream infections due to carbapenemase-producing Enterobacteriaceae (INCREMENT): A retrospective cohort study. Lancet Infect. Dis. 2017, 17, 726–734. [Google Scholar] [CrossRef]
- Motsch, J.; Murta de Oliveira, C.; Stus, V.; Köksal, I.; Lyulko, O.; Boucher, H.W.; Kaye, K.S.; File, T.M.; Brown, M.L.; Khan, I.; et al. RESTORE-IMI 1: A Multicenter, Randomized, Double-blind Trial Comparing Efficacy and Safety of Imipenem/Relebactam vs Colistin Plus Imipenem in Patients With Imipenem-nonsusceptible Bacterial Infections. Clin. Infect. Dis. 2020, 70, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Tumbarello, M.; Losito, A.R.; Giamarellou, H. Optimizing Therapy in Carbapenem-Resistant Enterobacteriaceae Infections. Curr. Opin. Infect. Dis. 2018, 31, 566–577. [Google Scholar] [CrossRef]
- Zhanel, G.G.; Golden, A.R.; Zelenitsky, S.; Wiebe, K.; Lawrence, C.K.; Adam, H.J.; Idowu, T.; Domalaon, R.; Schweizer, F.; Zhanel, M.A.; et al. Cefiderocol: A Siderophore Cephalosporin with Activity Against Carbapenem-Resistant and Multidrug-Resistant Gram-Negative Bacilli. Drugs 2019, 79, 271–289. [Google Scholar] [CrossRef]
- Otter, J.A.; Burgess, P.; Davies, F.; Mookerjee, S.; Singleton, J.; Gilchrist, M.; Parsons, D.; Brannigan, E.T.; Robotham, J.; Holmes, A.H. Counting the cost of an outbreak of carbapenemaseproducing Enterobacteriaceae: An economic evaluation from a hospital perspective. Clin. Microbiol. Infect. 2017, 23, 188–196. [Google Scholar]
- Jia, H.; Li, W.; Hou, T.; Ma, H.; Yang, Y.; Wu, A.; Liu, Y.; Wen, J.; Yang, H.; Luo, X.; et al. The Attributable Direct Medical Cost of Healthcare Associated Infection Caused by Multidrug Resistance Organisms in 68 Hospitals of China. BioMed Res. Int. 2019, 2019, 7634528. [Google Scholar] [CrossRef] [Green Version]



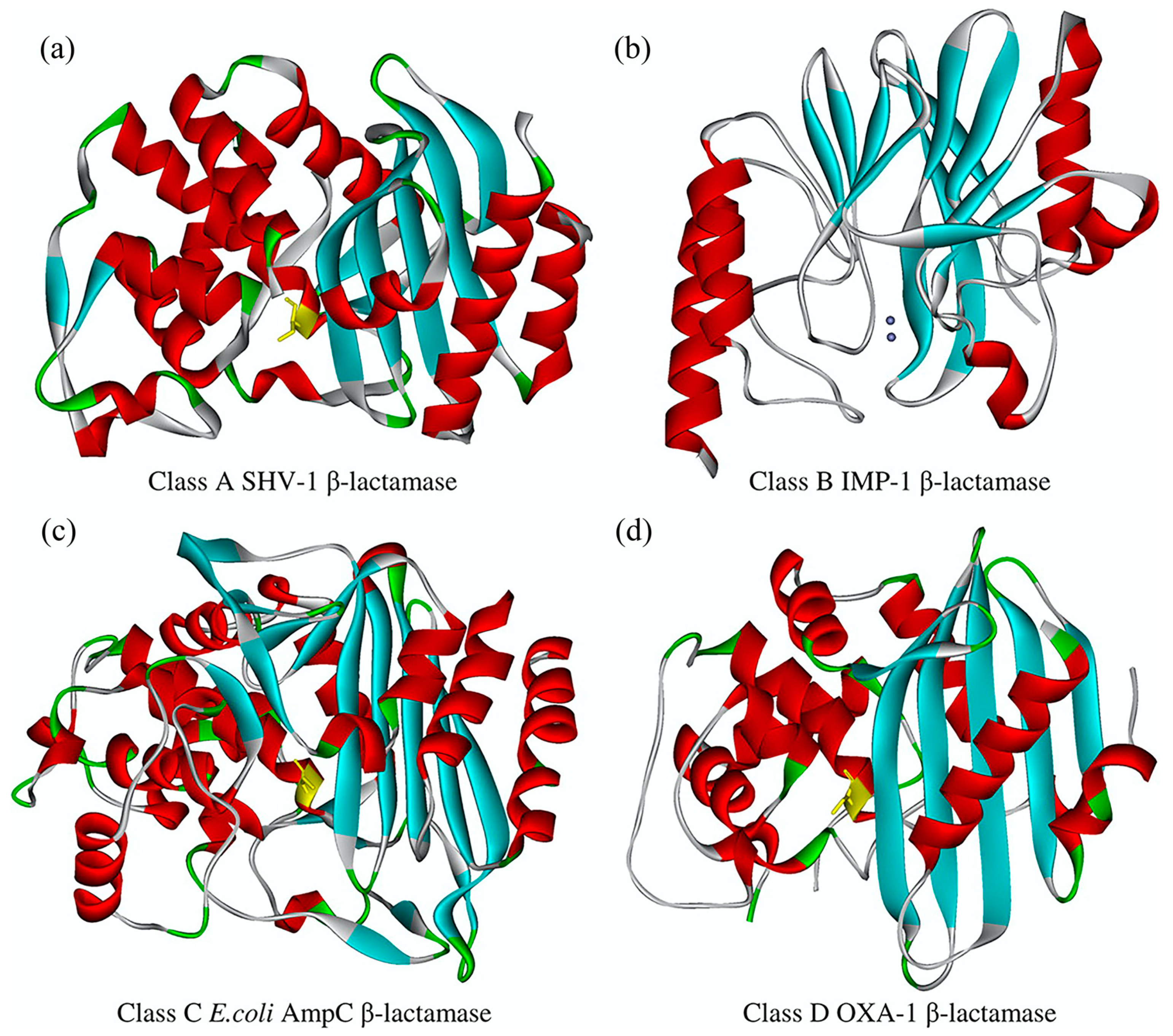{kind=link}
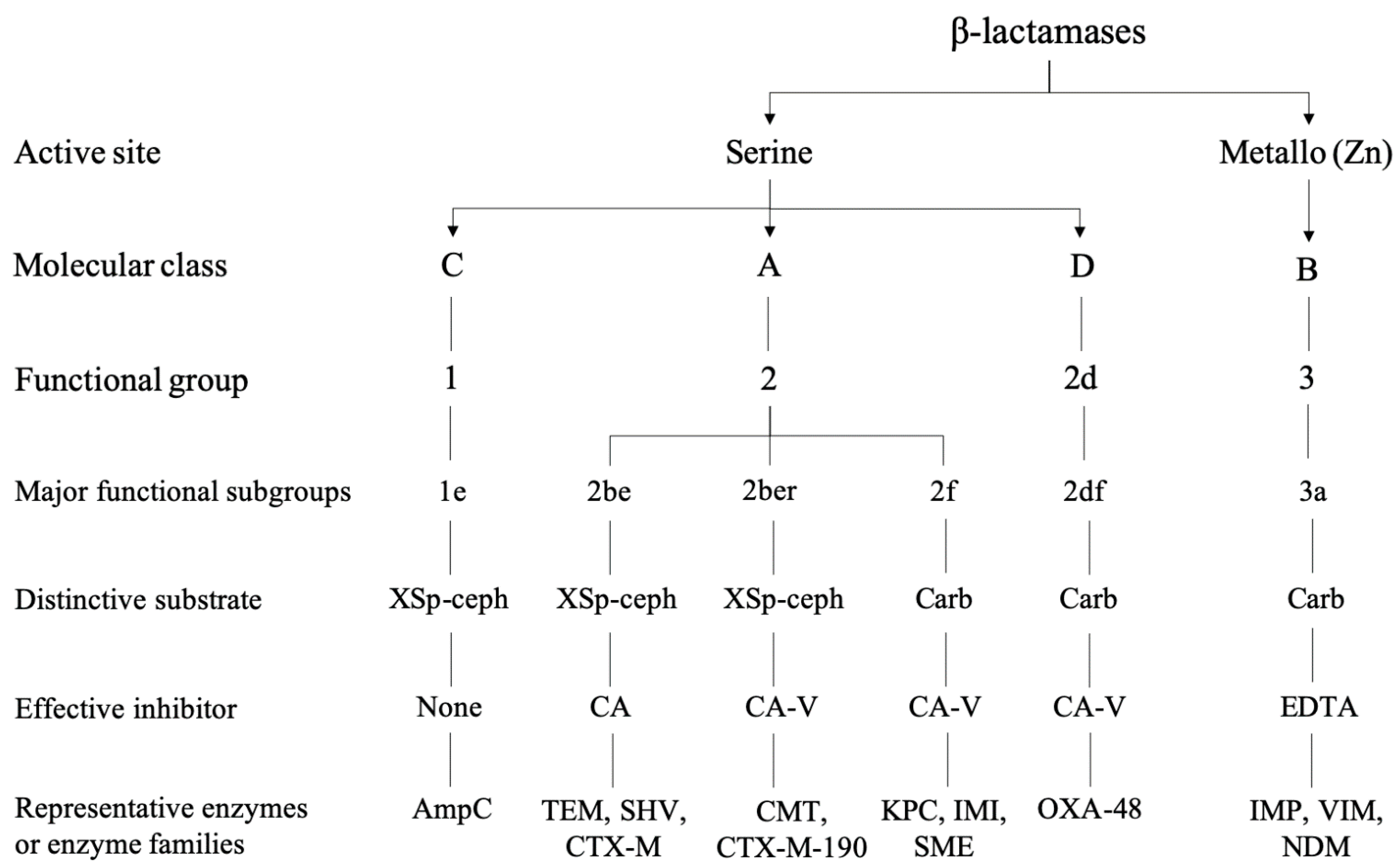{kind=link}
| Original (Current) Name | Microorganism | Year (Country) of First Isolation |
|---|---|---|
| TEM-1 | Escherichia coli | 1963 (Greece) |
| SHV-1 | Klebsiella pneumoniae | 1972 (Unknown) |
| Transferable ESBL (SHV-2) | Klebsiella pneumoniae | 1983 (Germany) |
| Serine (class A, group 2f) | Serratia marcescens | 1982 (England) |
| Carbapenemase (SME-1) | Serratia marcescens | 1985 (United States) |
| FEC-1 (CTX-M-1) | Escherichia coli | 1986 (Japan) |
| Plasmid-encoded AmpC (MIR-1) | Klebsiella pneumoniae | 1988 (United States) |
| Plasmid-encoded MBL (IMP-1) | Serratia marcescens | 1988 (Japan) |
| Inhibitor-resistant TEM (TEM-30) | Escherichia coli | 1991 (France) |
| KPC-type (KPC-2) | Klebsiella pneumoniae | 1996 (United States) |
| NDM-1 | Klebsiella pneumoniae | 2006 (India) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Angelis, G.; Del Giacomo, P.; Posteraro, B.; Sanguinetti, M.; Tumbarello, M. Molecular Mechanisms, Epidemiology, and Clinical Importance of β-Lactam Resistance in Enterobacteriaceae. Int. J. Mol. Sci. 2020, 21, 5090. https://doi.org/10.3390/ijms21145090
De Angelis G, Del Giacomo P, Posteraro B, Sanguinetti M, Tumbarello M. Molecular Mechanisms, Epidemiology, and Clinical Importance of β-Lactam Resistance in Enterobacteriaceae. International Journal of Molecular Sciences. 2020; 21(14):5090. https://doi.org/10.3390/ijms21145090
Chicago/Turabian StyleDe Angelis, Giulia, Paola Del Giacomo, Brunella Posteraro, Maurizio Sanguinetti, and Mario Tumbarello. 2020. "Molecular Mechanisms, Epidemiology, and Clinical Importance of β-Lactam Resistance in Enterobacteriaceae" International Journal of Molecular Sciences 21, no. 14: 5090. https://doi.org/10.3390/ijms21145090