Age, Sex Hormones, and Circadian Rhythm Regulate the Expression of Amyloid-Beta Scavengers at the Choroid Plexus


 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
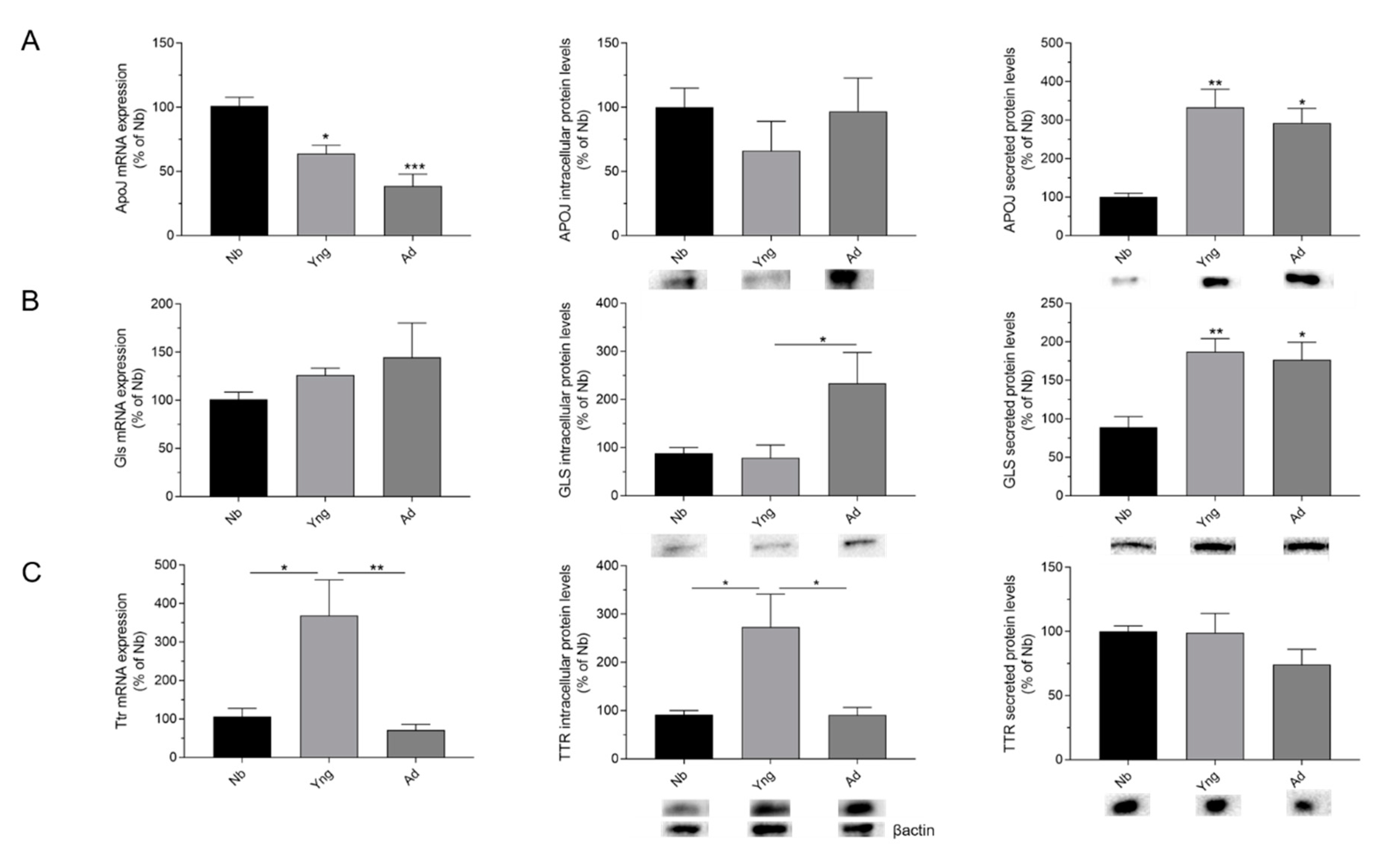
2.1. Effects of Age on the Expression of APOJ, GLS, and TTR in the Choroid Plexus
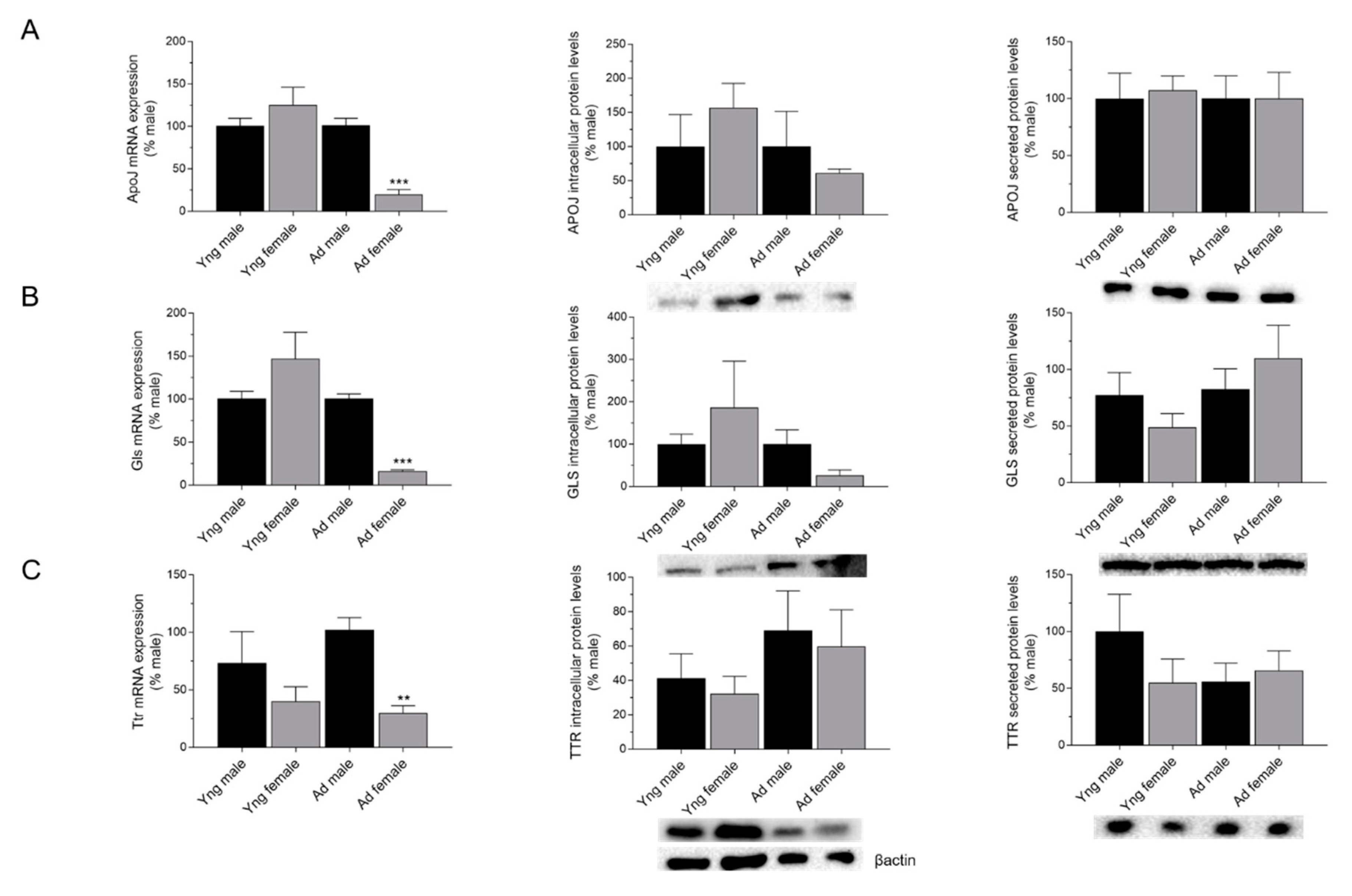
2.2. Effects of Sex on the Expression of APOJ, GLS, and TTR in the Choroid Plexus
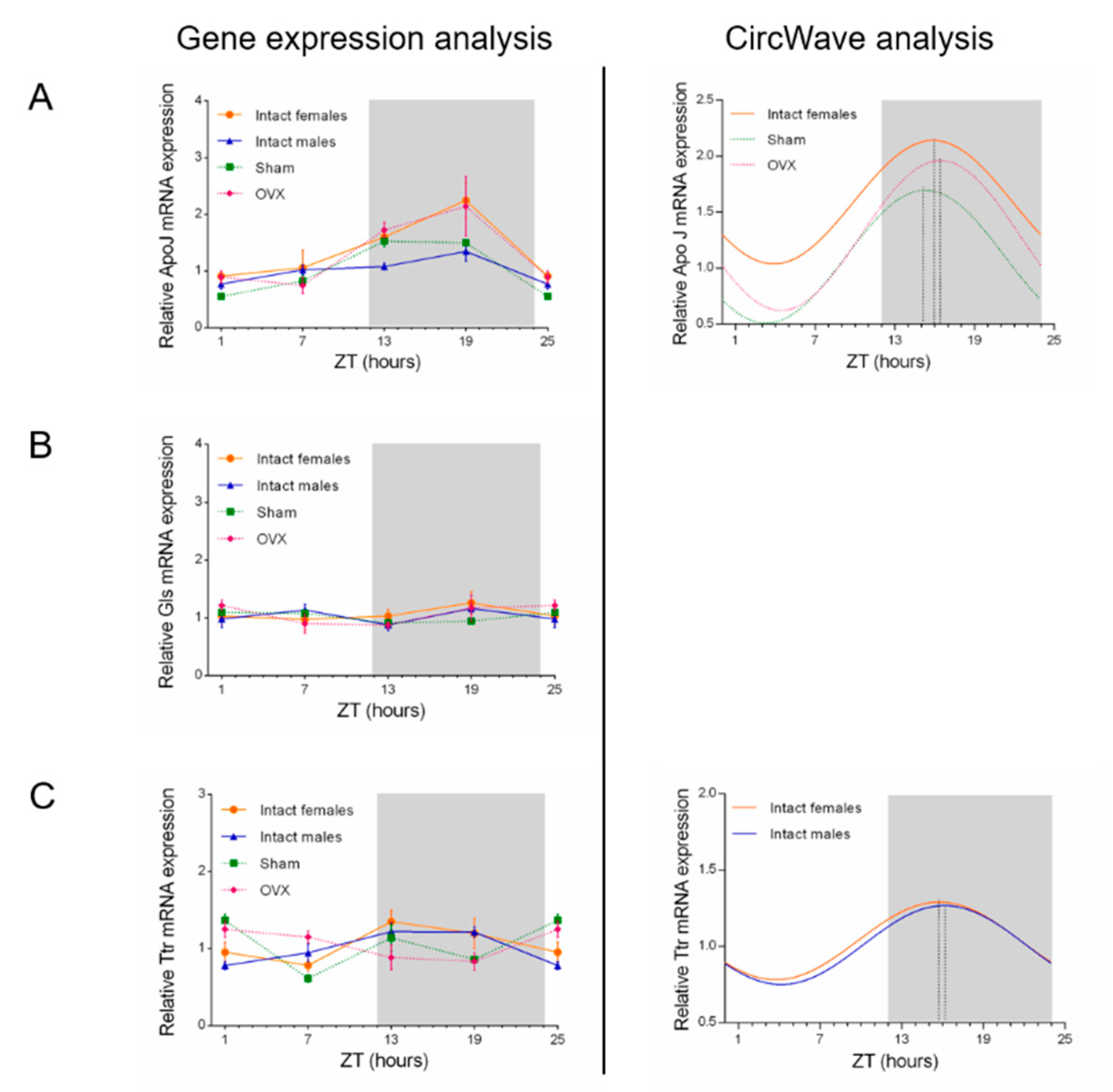
2.3. Influence of Circadian Rhythm in ApoJ, Gls, and Ttr Expression Levels
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Design—Effects of Age and Sex
4.3. Experimental Design—Circadian Effects
4.4. Real Time RT-PCR
4.5. Western Blot
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CP | Choroid plexus |
| Aβ | Amyloid beta |
| APOJ | Apolipoprotein J |
| GLS | Gelsolin |
| TTR | Transthyretin |
| CSF | Cerebrospinal fluid |
| BCSFB | Blood-cerebrospinal fluid barrier |
| AD | Alzheimer’s disease |
References
- Lehtinen, M.K.; Bjornsson, C.S.; Dymecki, S.M.; Gilbertson, R.J.; Holtzman, D.M.; Monuki, E.S. The choroid plexus and cerebrospinal fluid: Emerging roles in development, disease, and therapy. J. Neurosci. 2013, 33, 17553–17559. [Google Scholar] [CrossRef] [PubMed]
- Crossgrove, J.S.; Li, G.J.; Zheng, W. The choroid plexus removes beta-amyloid from brain cerebrospinal fluid. Exp. Biol. Med. 2005, 230, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Alvira-Botero, X.; Carro, E.M. Clearance of amyloid-beta peptide across the choroid plexus in Alzheimer’s disease. Curr. Aging Sci. 2010, 3, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Vargas, T.; Ugalde, C.; Spuch, C.; Antequera, D.; Moran, M.J.; Martin, M.A.; Ferrer, I.; Bermejo-Pareja, F.; Carro, E. Abeta accumulation in choroid plexus is associated with mitochondrial-induced apoptosis. Neurobiol. Aging 2010, 31, 1569–1581. [Google Scholar] [CrossRef]
- Dietrich, M.O.; Spuch, C.; Antequera, D.; Rodal, I.; de Yebenes, J.G.; Molina, J.A.; Bermejo, F.; Carro, E. Megalin mediates the transport of leptin across the blood-CSF barrier. Neurobiol. Aging 2008, 29, 902–912. [Google Scholar] [CrossRef]
- Krzyzanowska, A.; Carro, E. Pathological alteration in the choroid plexus of Alzheimer’s disease: Implication for new therapy approaches. Front. Pharmacol. 2012, 3, 75. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Marrero, I.; Gimenez-Llort, L.; Johanson, C.E.; Carmona-Calero, E.M.; Castaneyra-Ruiz, L.; Brito-Armas, J.M.; Castaneyra-Perdomo, A.; Castro-Fuentes, R. Choroid plexus dysfunction impairs beta-amyloid clearance in a triple transgenic mouse model of Alzheimer’s disease. Front. Cell Neurosci. 2015, 9, 17. [Google Scholar] [CrossRef] [Green Version]
- Duarte, A.C.; Hrynchak, M.V.; Goncalves, I.; Quintela, T.; Santos, C.R. Sex Hormone Decline and Amyloid beta Synthesis, Transport and Clearance in the Brain. J. Neuroendocrinol. 2016, 28. [Google Scholar] [CrossRef] [Green Version]
- Carro, E. Gelsolin as therapeutic target in Alzheimer’s disease. Expert Opin. Ther. Targets 2010, 14, 585–592. [Google Scholar] [CrossRef]
- Pascale, C.L.; Miller, M.C.; Chiu, C.; Boylan, M.; Caralopoulos, I.N.; Gonzalez, L.; Johanson, C.E.; Silverberg, G.D. Amyloid-beta transporter expression at the blood-CSF barrier is age-dependent. Fluids Barriers CNS 2011, 8, 21. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Kang, H.; Xu, Q.; Chen, M.J.; Liao, Y.; Thiyagarajan, M.; O’Donnell, J.; Christensen, D.J.; Nicholson, C.; Iliff, J.J.; et al. Sleep Drives Metabolite Clearance from the Adult Brain. Science 2013, 342, 373–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.E.; Lim, M.M.; Bateman, R.J.; Lee, J.J.; Smyth, L.P.; Cirrito, J.R.; Fujiki, N.; Nishino, S.; Holtzman, D.M. Amyloid-beta dynamics are regulated by orexin and the sleep-wake cycle. Science 2009, 326, 1005–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roh, J.H.; Huang, Y.; Bero, A.W.; Kasten, T.; Stewart, F.R.; Bateman, R.J.; Holtzman, D.M. Disruption of the sleep-wake cycle and diurnal fluctuation of beta-amyloid in mice with Alzheimer’s disease pathology. Sci. Transl. Med. 2012, 4, 150ra122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myung, J.; Schmal, C.; Hong, S.; Tsukizawa, Y.; Rose, P.; Zhang, Y.; Holtzman, M.J.; De Schutter, E.; Herzel, H.; Bordyugov, G.; et al. The choroid plexus is an important circadian clock component. Nat. Commun. 2018, 9, 1062. [Google Scholar] [CrossRef]
- Quintela, T.; Sousa, C.; Patriarca, F.M.; Gonçalves, I.; Santos, C.R.A. Gender associated circadian oscillations of the clock genes in rat choroid plexus. Brain Struct. Funct. 2014, 220, 1251–1262. [Google Scholar] [CrossRef]
- Quintela, T.; Albuquerque, T.; Lundkvist, G.; Belin, A.C.; Talhada, D.; Gonçalves, I.; Carro, E.; Santos, C.R. The choroid plexus harbors a circadian oscillator modulated by estrogens. Chronobiol. Int. 2017, 35, 270–279. [Google Scholar] [CrossRef]
- Barron, A.M.; Pike, C.J. Sex hormones, aging, and Alzheimer’s disease. Front. Biosci. 2012, 4, 976–997. [Google Scholar]
- Bell, M.R. Comparing Postnatal Development of Gonadal Hormones and Associated Social Behaviors in Rats, Mice, and Humans. Endocrinology 2018, 159, 2596–2613. [Google Scholar] [CrossRef]
- Dohler, K.D.; Wuttke, W. Changes with age in levels of serum gonadotropins, prolactin and gonadal steroids in prepubertal male and female rats. Endocrinology 1975, 97, 898–907. [Google Scholar] [CrossRef]
- Ahn, H.J.; Bae, J.; Lee, S.; Ko, J.E.; Yoon, S.; Kim, S.J.; Sakuragi, N. Differential expression of clusterin according to histological type of endometrial carcinoma. Gynecol. Oncol. 2008, 110, 222–229. [Google Scholar] [CrossRef]
- Won, Y.S.; Lee, S.J.; Yeo, S.G.; Park, D. Effects of Female Sex Hormones on Clusterin Expression and Paclitaxel Resistance in Endometrial Cancer Cell Lines. Int. J. Med Sci. 2011, 9, 86–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- E Vandenbroucke, R. A Hidden Epithelial Barrier in the Brain with a Central Role in Regulating Brain Homeostasis. Implications for Aging. Ann. Am. Thorac. Soc. 2016, 13, S407–S410. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Mao, Z.; Woody, S.K.; Brinton, R.D. Sex differences in metabolic aging of the brain: Insights into female susceptibility to Alzheimer’s disease. Neurobiol. Aging 2016, 42, 69–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Q.; Stepaniants, S.B.; Mao, M.; Weng, L.; Feetham, M.C.; Doyle, M.J.; Yi, E.C.; Dai, H.; Thorsson, V.; Eng, J.; et al. Integrated Genomic and Proteomic Analyses of Gene Expression in Mammalian Cells. Mol. Cell. Proteom. 2004, 3, 960–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayapal, K.P.; Philp, R.J.; Kok, Y.J.; Yap, M.G.; Sherman, D.H.; Griffin, T.J.; Hu, W.S. Uncovering genes with divergent mRNA-protein dynamics in Streptomyces coelicolor. PLoS ONE 2008, 3, e2097. [Google Scholar] [CrossRef]
- Huber, M.; Bahr, I.; Krätzschmar, J.R.; Becker, A.; Müller, E.-C.; Donner, P.; Pohlenz, H.-D.; Schneider, M.R.; Sommer, A. Comparison of Proteomic and Genomic Analyses of the Human Breast Cancer Cell Line T47D and the Antiestrogen-resistant Derivative T47D-r. Mol. Cell. Proteom. 2003, 3, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Koussounadis, A.; Langdon, S.P.; Um, I.H.; Harrison, D.J.; Smith, V.A. Relationship between differentially expressed mRNA and mRNA-protein correlations in a xenograft model system. Sci. Rep. 2015, 5, 10775. [Google Scholar] [CrossRef] [Green Version]
- Trougakos, I.P.; Gonos, E.S. Regulation of clusterin/apolipoprotein J, a functional homologue to the small heat shock proteins, by oxidative stress in ageing and age-related diseases. Free. Radic. Res. 2006, 40, 1324–1334. [Google Scholar] [CrossRef]
- Foster, E.M.; Dangla-Valls, A.; Lovestone, S.; Ribe, E.M.; Buckley, N.J. Clusterin in Alzheimer’s Disease: Mechanisms, Genetics, and Lessons From Other Pathologies. Front. Neurosci. 2019, 13, 164. [Google Scholar] [CrossRef] [Green Version]
- Orts-Sebastian, A.; Ludin, N.M.; Pawley, M.D.M.; Cheeseman, J.F.; Warman, G.R. Impact of anaesthesia on circadian rhythms and implications for laboratory experiments. Exp. Neurol. 2019, 311, 318–322. [Google Scholar] [CrossRef]
- Musiek, E.S.; Xiong, D.; Holtzman, D.M. Sleep, circadian rhythms, and the pathogenesis of Alzheimer disease. Exp. Mol. Med. 2015, 47, e148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mott, N.N.; Pinceti, E.; Rao, Y.S.; Przybycien-Szymanska, M.M.; Prins, S.A.; Shults, C.L.; Yang, X.; Glucksman, M.J.; Roberts, J.L.; Pak, T.R. Age-dependent Effects of 17beta-estradiol on the dynamics of estrogen receptor beta (ERbeta) protein-protein interactions in the ventral hippocampus. Mol. Cell Proteom. 2014, 13, 760–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serot, J.M.; Christmann, D.; Dubost, T.; Couturier, M. Cerebrospinal fluid transthyretin: Aging and late onset Alzheimer’s disease. J. Neurol. Neurosurg. Psychiatry 1997, 63, 506–508. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.L.; Athauda, S.B.P.; Kassem, N.A.; Zhang, Y.; Segal, M.B.; Preston, J.E. Decrease of Transthyretin Synthesis at the Blood-Cerebrospinal Fluid Barrier of Old Sheep. J. Gerontol. Ser. A Boil. Sci. Med Sci. 2005, 60, 852–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintela, T.; Goncalves, I.; Baltazar, G.; Alves, C.H.; Saraiva, M.J.; Santos, C.R. 17beta-estradiol induces transthyretin expression in murine choroid plexus via an oestrogen receptor dependent pathway. Cell. Mol Neurobiol. 2009, 29, 475–483. [Google Scholar] [CrossRef]
- Quintela, T.; Gonçalves, I.; Martinho, A.; Alves, C.H.; Saraiva, M.J.; Rocha, P.M.; Santos, C.R.A. Progesterone Enhances Transthyretin Expression in the Rat Choroid Plexus In Vitro and In Vivo via Progesterone Receptor. J. Mol. Neurosci. 2010, 44, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Quintela, T.; Alves, C.H.; Goncalves, I.; Baltazar, G.; Saraiva, M.J.; Santos, C.R. 5Alpha-dihydrotestosterone up-regulates transthyretin levels in mice and rat choroid plexus via an androgen receptor independent pathway. Brain Res. 2008, 1229, 18–26. [Google Scholar] [CrossRef]
- Martinho, A.; Santos, C.R.; Goncalves, I. A distal estrogen responsive element upstream the cap site of human transthyretin gene is an enhancer-like element upon ERalpha and/or ERbeta transactivation. Gene 2013, 527, 469–476. [Google Scholar] [CrossRef]
- Gião, T.; Saavedra, J.; Cotrina, E.; Quintana, J.R.; Llop, J.; Arsequell, G.; Cardoso, I. Undiscovered Roles for Transthyretin: From a Transporter Protein to a New Therapeutic Target for Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 2075. [Google Scholar] [CrossRef] [Green Version]
- Myung, J.; Wu, D.; Simonneaux, V.; Lane, T.J. Strong Circadian Rhythms in the Choroid Plexus: Implications for Sleep-Independent Brain Metabolite Clearance. J. Exp. Neurosci. 2018, 12. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Group | ||||
|---|---|---|---|---|
| Gene | Intact Females | Intact Males | Sham-Operated | Ovariectomized |
| ApoJ | p-value = 0.022 COG = 15.88 | NS | p-value = 0 COG = 15.3 | p-value = 0.013 COG = 16.43 |
| Gls | NS | NS | NS | NS |
| Ttr | p-value = 0.0225 COG = 15.76 | p-value = 0.0004 COG = 16.15 | NS | NS |
| Gene | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) |
|---|---|---|
| Apoj | CTGACCCAGCAGTACAACGA | AGCTTCACCACCACCTCAGT |
| Gls | GGTGCAGAGGCTCTTCCAGG | CTGCCGGAGCCACACCACTG |
| Ttr | GGACTGATATTTGCGTCTGAAGC | ACTTTCACGGCCACATCGAC |
| CycA | CAAGACTGAGTGGCTGGATGG | GCCCGCAAGTCAAAGAAATTAGAG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duarte, A.C.; Furtado, A.; Hrynchak, M.V.; Costa, A.R.; Talhada, D.; Gonçalves, I.; Lemos, M.C.; Quintela, T.; Santos, C.R.A. Age, Sex Hormones, and Circadian Rhythm Regulate the Expression of Amyloid-Beta Scavengers at the Choroid Plexus. Int. J. Mol. Sci. 2020, 21, 6813. https://doi.org/10.3390/ijms21186813
Duarte AC, Furtado A, Hrynchak MV, Costa AR, Talhada D, Gonçalves I, Lemos MC, Quintela T, Santos CRA. Age, Sex Hormones, and Circadian Rhythm Regulate the Expression of Amyloid-Beta Scavengers at the Choroid Plexus. International Journal of Molecular Sciences. 2020; 21(18):6813. https://doi.org/10.3390/ijms21186813
Chicago/Turabian StyleDuarte, Ana C., André Furtado, Mariya V. Hrynchak, Ana R. Costa, Daniela Talhada, Isabel Gonçalves, Manuel C. Lemos, Telma Quintela, and Cecília R.A. Santos. 2020. "Age, Sex Hormones, and Circadian Rhythm Regulate the Expression of Amyloid-Beta Scavengers at the Choroid Plexus" International Journal of Molecular Sciences 21, no. 18: 6813. https://doi.org/10.3390/ijms21186813