The Role of Extracellular Vesicles (EVs) in the Epigenetic Regulation of Bone Metabolism and Osteoporosis


1
Department of Women’s and Children’s Health, University of Padova, 35122 Padova, Italy
2
Research Laboratories, Department of Onco-hematology, Pediatric Hospital Bambino Gesù, 00146 Rome, Italy
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2020, 21(22), 8682; https://doi.org/10.3390/ijms21228682
Submission received: 17 September 2020
/
Revised: 8 November 2020
/
Accepted: 10 November 2020
/
Published: 17 November 2020
(This article belongs to the Special Issue Epigenetic, microRNA and Long Non-Coding RNA Roles in Osteoporosis)
Abstract
:Extracellular vesicles (EVs) are complex phospholipidic structures actively released by cells. EVs are recognized as powerful means of intercellular communication since they contain many signaling molecules (including lipids, proteins, and nucleic acids). In parallel, changes in epigenetic processes can lead to changes in gene function and finally lead to disease onset and progression. Recent breakthroughs have revealed the complex roles of non-coding RNAs (microRNAs (miRNAs) and long non-coding RNAs (lncRNAs)) in epigenetic regulation. Moreover, a substantial body of evidence demonstrates that non-coding RNAs can be shuttled among the cells and tissues via EVs, allowing non-coding RNAs to reach distant cells and exert systemic effects. Resident bone cells, including osteoclasts, osteoblasts, osteocytes, and endothelial cells, are tightly regulated by non-coding RNAs, and many of them can be exported from the cells to neighboring ones through EVs, triggering pathological conditions. For these reasons, researchers have also started to exploit EVs as a theranostic tool to address osteoporosis. In this review, we summarize some recent findings regarding the EVs’ involvement in the fine regulation of non-coding RNAs in the context of bone metabolism and osteoporosis.
Keywords:
epigenetics; long non-coding RNA; microRNA; osteoblast; osteoclast; osteocyte; osteoporosis; bone metabolism1. Introduction
Osteoporosis is defined by the World Health Organization (WHO) as a progressive systemic skeletal disorder with low bone density and deterioration of bone architecture leading to an increased risk of bone fragility and fracture [1,2]. Currently, osteoporosis is a major public health problem. Although osteoporosis is not a severe disease per se, the healthcare cost is massive, especially for the management of the fractures resulting from low bone mass, associated with a decreased quality of life and lifespan in aging people. About 200 million people (1 in 8 women after 55 years of age and 1 in 4 men after 65 years of age) suffer from osteoporosis worldwide with an associated incidence of 9 million fractures [3,4]. The problem is growing in an aging population. In women, the increase in the bone remodeling rate in both cortical and cancellous bone, combined with a negative remodeling balance, results in bone loss and collapse of bone microarchitecture. Indeed, cortical bone peculiarly shows an increase in cortical porosity and a reduction in cortical thickness, while cancellous bone is mainly affected by thinning and loss of trabeculae [5,6,7]. In men, a reduction in bone formation and bone turnover occurs during aging. Changes in matrix and mineral composition of bone can also contribute to increased bone fragility [8]. It has been shown that osteoporosis has genetic and molecular heterogeneity like many other common complex diseases [3]. The era of genome-wide association studies (GWAS) clarifies the horizon of the genetic contribution to osteoporosis. Many GWAS have been performed interrogating genetic association with bone mineral density (BMD), implicating more than 90 candidate genes for osteoporosis [9]. In this context, non-coding RNAs were identified as playing a crucial role as a genomic regulatory system in several biological processes of bone metabolism [10,11]. Interestingly, some GWAS reported association between polymorphisms of ncRNAs and BMD. Zeng et al. conducted a meta-analysis among individuals of seven independent studies for BMD, reporting that MEF2C antisense RNA 1 (MEF2C-AS1) was significantly associated with BMD of femoral neck, while LOC100506136 showed significant association with BMD of hip [12]. Similarly, Zhang et al. constructed a comprehensive mRNA–lncRNA–miRNA competing endogenous RNA (ceRNA) regulatory network to predict the risk of osteoporosis development by analyzing public Gene Expression Omnibus (GEO) microarray profile data [13]. In the next paragraphs, we provide information about the mechanisms and the role of non-coding RNAs in the regulation of gene expression and their link with extracellular vesicles (EVs) in the onset of osteoporosis.
1.1. Cellular Basis of Bone Metabolism and Osteoporosis
Osteoporosis occurs when the bone turnover is perturbed and bone mass decreases [1]. Bone mass is maintained by the synchronized and fine-tuned crosstalk between bone building cells, osteoblasts, and bone-resorbing cells, i.e., osteoclasts. On the one hand, osteoblasts, deriving from mesenchymal precursors, are responsible for the deposition of both the organic (mainly collagen type I) and inorganic (hydroxyapatite Ca10(PO4)6(OH)2) constituents of the extracellular matrix [14]. On the other hand, osteoclasts, multinucleated giant cells arising from the fusion of mononuclear hematopoietic precursors of the monocytic/macrophagic lineage, degrade the extracellular matrix through a very specialized and unique structure, the resorption lacuna, which allows the hydrolysis of hydroxyapatite through the acidification of extracellular space and the subsequent exposition of collagen fibrils, degraded by metalloproteases and cathepsin K [15,16]. The activity of these cells is strictly coordinated in time and space, and the interdependence of these cells is such that they are grouped into one histological and functional unit, called the bone remodeling unit [17,18]. However, these two cell types are not the only players on the stage [18]. In fact, the most abundant (>90%) cells in the bone are the osteocytes, deriving from the embedding in the bone matrix of mature osteoblasts [19]. Osteocytes, previously considered static bystanders entrapped into the bone, are now known to play a crucial role in sensing mechanical loading and regulating calcium and phosphate homeostasis [20]. These signals are integrated and reported to the bone remodeling unit, since osteocytes create a tight web across the skeleton with the projection of 40–100 dendritic extensions per cell, which spread through the bone matrix in channels called canaliculi, forming connections to neighboring osteocytes, osteoblasts, osteoclasts, and blood vessels. Many molecular mechanisms concur to synchronize the osteoblast/osteoclast/osteocyte activity. The main one is the receptor activator of NF-κB ligand, RANKL [15,21]. This cytokine, belonging to the tumor necrosis factor (TNF) superfamily members, is expressed on the surface of osteoblasts and osteocytes. The binding of RANKL to its receptor RANK on the membrane of osteoclast precursors and mature osteoclasts triggers the osteoclast formation and sustains the osteoclast function. This ligand–receptor binding can be prevented by osteoprotegerin (OPG), a soluble decoy of RANKL, produced by osteoblasts and osteocytes. In addition, osteocytes influence in a paracrine manner both osteoblast and osteoclast activity, i.e., osteocytes regulate bone formation by secreting modulators of the wingless-type mouse mammary tumour virus [MMTV] integration site members (Wnt) signaling pathway such as activators (nitric oxide and ATP) and inhibitors (sclerostin (SOST) and dickkopf-related protein 1 (DKK1)). Similarly, osteocytes regulate bone resorption, other than by RANKL and OPG, by releasing nitric oxide, an inhibitor of osteoclast formation and activity [22]. The amounts of bone resorbed and formed in each bone remodeling unit are important determinants of bone mass. This continuous bone remodeling process is required to replace old bone tissue and to repair bone micro-cracks [17]. In adults, it has been estimated that 10% of bone is replaced every year and that this is crucial for the maintenance of a healthy skeleton [17].
Many determinants, both genetic and environmental, contribute to set bone mass and guarantee bone health. Steroid hormones exert a crucial role in reaching (during puberty) and maintaining (at adulthood) the peak of bone mass and the correct BMD level among both women and men [23]. In fact, the decline of estrogen and testosterone during menopause and andropause is one of the major risk factors and drivers of osteoporosis [23,24]. Hormonal withdrawal also contributes to mineral disturbances. Indeed, intestinal calcium absorption is decreased and urinary calcium excretion is increased, thus leading to a negative calcium balance and secondary hyperparathyroidism, exacerbating osteoclast activity [25,26]. Physical exercise and muscle fitness have a dramatic impact on bone health. In fact, the mechanical force produced during muscular activity is sensed by osteocytes and converted into bone deposition, whereas disuse and/or muscle atrophy result in osteoporosis [27,28]. Moreover, muscle secretes a set of molecules, known as myokines, directly affecting bone metabolism, such as irisin, myostatin, and insulin-like growth factor-1 [29].
1.2. Diagnosis and Biomarkers of Osteoporosis
The standard clinical technique for diagnosis of osteoporosis is the dual-energy x-ray absorptiometry (DXA) of the spine or hip [21]. DXA reports the T-score, a quantified measure of BMD. A T-score between −1 and −2.4 standard deviations compared to the mean score of a healthy young woman defines an osteopenic condition, whereas a T-score greater than or equal to −2.5 standard deviations is considered diagnostic for osteoporosis [22]. BMD values are also routinely used to monitor fracture risk and bone health. However, BMD has several limitations, i.e., it is not effective and sensitive as a surveillance tool, especially in the case of minimal changes during a pharmacological treatment, and the T-score is not in many cases a reliable predictor of fragility fractures. Due to these limitations, the management of osteoporosis in most cases mainly relies on biomarkers [30,31]. These are mostly molecules released by bone cells during bone remodeling, measurable in urine or serum, and are indicative of the rate of bone turnover. Bone biomarkers can be classified into markers of bone formation or bone resorption [32]. The first class groups total alkaline phosphatase (total ALP), bone-specific alkaline phosphatase (B-ALP), procollagen type 1 N-terminal propeptide (P1NP), osteocalcin (OC), and procollagen type 1 C-terminal propeptide (P1CP). The second class includes hydroxyproline (HYP), pyridinoline, tartrate-resistant acid phosphatase 5b (TRAP 5b), deoxypyridinoline (DPD), carboxy-terminal cross-linked telopeptide of type 1 collagen (CTX-1), and amino-terminal cross-linked telopeptide of type 1 collagen (NTX-1).
1.3. Drugs and Therapeutic Strategies for Osteoporosis Management
Pharmacological opportunities for osteoporosis aim to restore the steady activity of the bone remodeling unit. Bone anti-resorptive and bone anabolic agents are available in clinical management [32].
1.3.1. Anti-Resorptive Agents
Anti-resorptive drugs are the most commonly used pharmacological tool to avoid osteoporosis progression. These drugs abrogate the exacerbated osteoclast activity, preserving the existing bone mass and structure. Bisphosphonates (alendronate, risedronate, and zoledronic acid) are the first-in-use drugs used to inhibit bone resorption and, to date, are still largely used in the clinical management of osteoporosis [30,33]. The newest agent denosumab, a fully human monoclonal antibody, binds to and inhibits RANKL, resulting in a marked but reversible inhibition of bone resorption.
1.3.2. Anabolic Agents
Anabolic agents aim to improve osteoblast function and stimulate bone deposition and mineralization. Estrogen-replacing therapy prevents bone loss in postmenopausal women [34]. Use is limited in early menopause, no more than 10 years, because of concerns about cardiovascular safety [35]. Selective estrogen receptor modulators (SERMs, such as raloxifene and bazedoxifene) bind with high affinity to the estrogen receptor and have both agonist and antagonist properties depending upon the target tissue [36]. SERMs present the same procoagulant concerns of estrogen, but exert a beneficial effect to reduce breast cancer [37]. Parathyroid hormone (PTH) receptor agonists, e.g., teriparatide (recombinant fragment of human PTH consisting of the first 34 amino acids) and abaloparatide (analogue of PTH-related peptide, PTHrP) improve bone mass and reduce fracture through binding of the PTH receptor-1 on osteoblasts [38,39]. The newest FDA-approved anabolic agent is romosozumab, a monoclonal antibody that binds to SOST [40]. Due to the inhibition of Sost, romosozumab both increases bone formation and decreases bone resorption.
2. Epigenetic Regulation
Epigenetics is the study of inherited changes in phenotype or gene expression that are caused by mechanisms other than changes in the structural DNA sequence [41,42]. These modifications may persist through several cell divisions lasting for generations or can be acquired permanently. The most noteworthy epigenetic mechanisms include the following: (1) DNA methylation, through which the genome is locally regulated inserting a methyl group tag on DNA by DNA methyltransferases; (2) histone modifications, through which the nucleosome superstructure is regulated by reversibly modulating the DNA wrapping extent around histones by histone methyltransferases and histone acetyltransferases; and (3) processes mediated by the most recently discovered class of RNAs, the non-coding RNAs [43]. Although actively transcribed, these RNAs are not translated into proteins and can be generally divided into two categories based on size, namely short-chain non-coding RNAs (sncRNAs) and long non-coding RNA (lncRNAs) [43,44,45]. A better ncRNA classification relies on the structure or function. microRNAs (miRNAs), which are 19–24 nucleotides in length, usually recognize target mRNAs by complementarity to a 2–7 nucleotide long seed region in the 3′-UTR region, although the seed region can be also located at the 5′-UTR and CDS regions. Their biogenesis is dependent on Drosha, Dicer, and Argonautes (AGO) and, finally, activating RNA-induced silencing (RISC) complex [46,47]. The Piwi-interacting RNAs (piRNAs), are 26–30 nucleotide single-stranded RNAs, are Dicer independent, and interact with Piwi-domain-containing proteins [48]. The piRNA precursor transcripts undergo endonucleolytic cleavage in the cytoplasm and are complexed with the Aubergine (Aub) or Piwi chaperone complex for 3′ trimming and methylation, finalizing the primary biogenesis pathway. Natural small interfering RNAs (siRNAs) are 20 to 24 base pair double-stranded RNAs (dsRNAs) with phosphorylated 5′ ends and hydroxylated 3′ ends with two overhanging nucleotides [43,49]. Upon Dicer cleavage, a shorter double-strand duplex is formed ready to enter into the RISC. Small nuclear RNAs (snRNAs) and small nucleolar RNAs (snoRNAs), are approximately 150 nucleotide transcripts found within the nucleus of eukaryotic cells, involved in the processing of pre-messenger RNA, in the regulation of transcription factor, and in the maintaining of telomeres, or in ribosomal RNA modification and processing in the nucleolus [48]. Finally, the long non-coding RNAs can be classified as follows, according to their structural relationship with the adjacent protein-coding genes: (a) sense or antisense, lncRNAs are located and transcribed on the same or the opposite strand of the adjacent protein-coding genes; (b) convergent or divergent, lncRNAs have a convergent (divergent) orientation of transcription compared with that of the adjacent protein-coding genes; and (c) intergenic or intronic, lncRNAs are located between two protein-coding genes, or located in introns [45,50,51,52,53]. The most abundant isoform of lncRNAs are predicted to be circular RNAs (circRNAs) compared to a linear transcript. CirclncRNAs can be formed via canonic splicing (intronic circRNAs) or via backsplicing, joining the splice donor at the upstream acceptor site [54,55,56]. Another classification of lncRNAs is relative to their functions, as discussed below (see Section 2.2).
In recent years, a large number of studies have shown that non-coding RNAs play a significant role in epigenetic modifications at post-transcriptional or promoter levels, since they can regulate gene expression both at the level of the gene itself and at the level of the chromosome to control cell differentiation [57,58,59].
2.1. miRNAs
miRNAs are single-stranded RNAs of approximately 19–24 nucleotides, of which 50% are located in non-coding chromosomal regions [60]. During biogenesis (Figure 1), a primordial hairpin-shaped double-stranded intermediate is maturated by Drosha and DICER enzymes [61]. Finally, the mature miRNA is complexed with the loading proteins AGO, being able to target a specific messenger RNA through the assembly with RISC [46]. Almost 1800 putative miRNAs have been identified in the human genome, and the number of miRNAs is still increasing rapidly due to the progress in sequencing and bioinformatic technologies [62]. Interestingly, a single miRNA can target up to hundreds of different genes [63]. Histone methyltransferases, methyl CpG-binding proteins, chromatin domain proteins, and histone deacetylases are assumed to be putative targets for miRNAs [63]. For example, histone deacetylase 4 (HDAC4) has been demonstrated to be a specific target for miR-140 in mouse embryonic cartilage tissue [64]. miRNAs (i.e., the miR-29 family) can also regulate DNA methylases, affecting DNA methyltransferases 3A and 3B [65].
miRNA dysfunctions have been associated with different pathologies, including osteoporosis. Three miRNAs (miR-21, miR-133a, and miR-146a) have been found differentially expressed in the plasma of 120 Chinese postmenopausal women according to their T-scores [20]. In the serum of osteoporotic patients, Seelinger and colleagues identified five overexpressed miRNAs (miR-100, miR-122, miR-124a, miR-125b and miR-148a) associated with a high fracture risk [66]. Along the same lines, another group confirmed the differential expression of five miRNAs (miR-10a/b, miR-133b, miR-21-5p, and miR-22-3p) in the serum of postmenopausal women suffering from recent femoral-neck fractures [67].
2.2. Long Non-Coding RNAs (lncRNAs)
lncRNAs are a newly discovered class of non-coding regulatory RNAs generally containing more than 200 nucleotides. They are located in the nucleus or cytoplasm, and rarely encode proteins [68]. lncRNAs can be generated by the following five main mechanisms: (a) disruption of the translational reading frame of a gene, (b) chromosomal reorganization, (c) retrotransposition of a non-coding gene, (d) partial tandem duplication of a non-coding gene, and (e) insertion of a transposable element into a gene producing a functional non-coding gene [45]. lncRNAs can behave as signal, favoring gene expression; decoy, suppressing gene expression; guide, promoting chromatin modification; or scaffold, acting on a chromatin structure [69]. Furthermore, lncRNAs can work as a sponge for miRNAs, since they can harbor recognition sites for miRNAs. Due to the complementarity of the sequences, lncRNAs can sequester miRNA and prevent it from repressing its mRNA target, resulting in increased expression of protein [70]. In this view, lncRNA H19 has been reported to enhance osteoblast proliferation by sequestering miR-141 and miR-22, negative regulators of osteogenesis and the Wnt/β-catenin pathway [71]. Similarly, lncRNA metastasis-associated lung adenocarcinoma transcript 1 (MALAT 1) acted as miR-1 sponge to inhibit its suppressive transcription effect on Connexin43 [72]. For these reasons, these lncRNAs were defined as competing endogenous RNAs (ceRNAs) (Figure 1). lncRNAs have been associated with the development of osteoporosis, and some authors proposed their use as a diagnostic and therapeutic tool for osteoporosis [73,74,75,76,77,78]. For example, X-inactive-specific transcript (Xist) RNA has been described to inhibit osteogenic differentiation bone marrow mesenchymal stem cell [79]. Interestingly, Xist was associated with the inhibition of the osteogenic potential of bone marrow mesenchymal stem cells, suppressing protein expression levels of ALP, OC, and runt-related transcription factor 2 (RUNX2) [79]. Accordingly, Xist was highly expressed in patients affected with osteoporosis [79]. Along the same lines, lncRNA ZBTB40-IT1 has been revealed as a regulator of bone metabolism, since it is able to suppress osteogenesis and promote osteoclastogenesis by regulating the expression of WNT4, RUNX2, Osterix (OSX), ALP, collagen type I alpha 1 chain (COL1A1), OPG, and RANKL in osteoblastic-like cells [80]. Similarly, low expression of lncRNA H19 has been detected in disuse osteoporosis, accompanied by the inhibition of osteogenesis and impaired trabecular bone growth [81].
3. Extracellular Vesicles
Currently, the intricate scenario of cell-to-cell communication is further complicated by the recognition of the pivotal role of EVs. EVs are complex phospholipidic cell-derived structures actively released from cells in the environment and able to shuttle biological information across cells and tissues. Although the involvement of EVs in virtually all biological processes (i.e., embryogenesis, neuronal plasticity, immune response) was demonstrated in the last decade, the existence of EVs was reported by Wolf in 1967, associated with platelets and coagulation [82], and they were subsequently observed in reticulocyte differentiation, acting as a cellular garbage system [83]. Since then, extensive research activity in the field has led to a better—although largely incomplete—understanding of their biological properties and has laid the foundation for their use as diagnostic and therapeutic tools [84]. Extracellular vesicles are classified into the following three main classes, accordingly to Minimum Information for Studies of EVs (MISEV) guidelines: small extracellular vesicles (sEVs, diameter < 200 nm, also known as exosomes), medium/large extracellular vesicles (m/lEVs, diameter >200 nm up to 1000 nm, also known as microvesicles), and apoptotic bodies (>1000 nm up to 5000 nm) [85]. Apart from the size, the three classes of EVs differ in their biogenesis [86] (Figure 2). Small EVs derive from the release of a larger structure, called a multivesicular body (MVB), a component of the endocytic pathway that sorts sEVs in its lumen, finally releasing them by the fusion with the plasma membrane. This mechanism involves multiple protein partners, such as Ras-related proteins in brain (RAB), endosomal sorting complex required for transport (ESCTR) components, and proteins in the ceramide/sphingomyelinase pathway [86]. Medium/large shedding EVs, or microvesicles, bud from the cell surface by sprouting and scission of the membrane. ADP-ribosylation factor 6 (ARF6) activates the phospholipase D (PLD), resulting in a phospholipidic reorganization and thereby relocating phosphatidylserines toward the outer side of the membrane. Finally, extracellular signal-regulated kinase (ERK) is recruited to the plasma membrane and activates through phosphorylation the myosin light-chain kinase (MLCK), resulting in invagination of plasma membrane and release of EVs [86]. Apoptotic bodies arise from the cleavage of the cell during apoptosis, a way to neatly package the cell components in the surroundings and exert many biological effects [87,88].
Interestingly, all the resident bone cells, i.e., osteoblasts, monocytes/macrophages, osteoclasts, osteocytes, adipocytes, and endothelial cells, have been demonstrated to release EVs or respond to EVs both in physiological and pathological conditions [89,90,91,92,93,94,95,96]. EVs have been identified to shuttle molecules coming from the molecular legacy of the donor cells, exerting a specific effect according to the metabolic status of parental cells [97]. A multitude of studies defined the involvement of EVs in transferring genetic materials in many systems. For example, monocyte-derived EVs shuttle miR-155 to the endothelium, increasing endothelial cell migration [98]. Lv et al. demonstrated that vesicular tubular epithelial cells communicate by EVs shuttling miR-19b-3p to macrophages, leading to M1 macrophage switching [99]. Multiple myeloma cells under hypoxic conditions release miR-135b enriched sEVs able to decrease in target endothelial cells the factor-inhibiting hypoxia-inducible factor 1 (FIH-1), thereby increasing angiogenesis [100]. For a more detailed review on this aspect, see O’Brien et al. [101].
4. Control of Bone Metabolism by Means of EVs
Communication by means of EVs is a crucial mechanism involved in bone metabolism and intercellular crosstalk [14]. Some evidence in the late 1960s prompted researchers to postulate a role played by EVs in early mineral nucleation during cartilage mineralization [102,103]. More recently, Davies et al. demonstrated that mineralizing osteoblasts release EVs enriched in annexin 2, making the EV membrane able to complex octacalcium phosphate and other ions. This complex showed the intrinsic ability of triggering mineralization in an acellular context [104]. Furthermore, EVs are strictly related to cell-to-cell communication. Osteoclast differentiation and survival required the action of the irreplaceable cytokine receptor activator of NF-κB ligand (RANKL) [105,106,107]. Different groups described the release from osteoblast-like cells of EVs enriched in RANKL, directly supporting osteoclastogenesis [89,108]. In contrast, Ikebuchi et al. described that the mature osteoclasts produce EVs shuttling RANK [91]. Once bound to RANKL expressed on the osteoblast membrane, EVs trigger the reverse signaling pathway, inducing osteoblast maturation and bone deposition. Weilner and colleagues showed that endothelial cells produce EVs containing galectin-3, able to induce osteogenic differentiation on MSCs [109]. Adipocytes transfer via EVs adipocyte-specific transcripts such as adiponectin, resistin, and peroxisome proliferator-activated receptor gamma 2 (Pparγ2) into macrophages [110] or leptin, tumor necrosis factor alpha (Tnfα), and fibroblast growth factor alpha (Fgfα) into endothelial cells, thereby inducing angiogenesis [111]. Finally, peripheral blood mononuclear cells transfer via EVs the chemokine receptor CCR5 to endothelial cells [112]. Endothelial precursors have been described to secrete EVs able to attenuate steroid-induced osteoblast apoptosis and autophagy, being able to upregulate glutathione peroxidase 4, system Xc−, and cysteine levels while reducing malondialdehyde and reactive oxygen species production [113].
Here, we discuss in more detail some examples of miRNA and lncRNA transfer by EVs among the bone cells, focusing on mechanisms detrimental to bone quality (Figure 3).
5. Epigenetic Detrimental Effects on Bone Metabolism by Means of EVs
5.1. Mesenchymal-Derived EVs
Human bone marrow stromal cells (hBMSCs) release EVs enriched in miRNAs, whose profile is different according to the metabolic status of the cells [114]. Among the identified miRNAs, Xu et al. characterized miR-885-5p. This miRNA was found inside EVs and its expression decreased during osteogenic differentiation of hBMSCs. Overexpression of miR-885-5p in hBMSCs impaired wnt5a and runx2, finally dampening their osteogenic potential.
Osteoblast-like cells MC3T3-E1 have been demonstrated to release EVs affecting mineralization of bone marrow stromal cell line ST2. These EVs were enriched in many miRNAs, among which miR-30d-5p, miR-133b, and miR-140-3p are reported [115]. A previous study already demonstrated that miR-30d-5p and miR-133b inhibit osteoblast differentiation targeting RUNX2, the master gene of osteoblast differentiation [116].
Different studies highlighted the association and contribution of miR-146a to the progression of different degenerative diseases, such as rheumatoid arthritis, systemic lupus erythematosus, and myelodysplastic syndrome, as well as osteoporosis, both in rodents and in patients [20,117]. Zhao et al. demonstrated that miR-146a supports RANKL expression in osteoblasts, enhancing osteoclast function and bone turnover, showing that miR-146a−/− mice are protected from ovariectomy (OVX)-induced osteoporosis [117]. Cao and colleagues noted the TNFα-induced cell growth arrest of osteoblast-like MC3T3 cells by the upregulation of miR-146a [118]. Interestingly, immune cells are reported to secrete miR-146a-enriched sEVs [119]. Finally, Dong et al. reported that exosomes derived from the serum of systemic lupus erythematosus patients are enriched in miR-146a and induced the senescence of MSCs [120].
MSCs differentiate towards osteogenic or adipogenic precursors, and the osteoblast/adipocyte balance is tightly regulated at the level of gene transcription [121]. Adipogenesis is known to negatively affect osteogenesis and osteoblast function, and EVs from adipocytes participate in the inhibition of osteoblast differentiation [122]. In fact, MSCs treated with adipocytic EVs increased the transcriptional level of PPARγ, leptin, CEBPα, and CEBPδ, whereas they decreased osteocalcin and osteopontin levels [123]. These effects were ascribable to miR-138, miR-30c, miR-125a, miR-125b, and miR-31 miRNAs detectable in the adipocytic EVs, and in the MCSs upon incubation with adipocytic EVs, confirming a horizontal transfer of genetic material between these cells.
5.2. Immune Cell- and Osteoclast-Derived EVs
Ma et al. described that mononuclear cells and granulocytes release EVs shuttling the lncRNA JPX under some pathological circumstances in women [124]. Interestingly, JPX is known to be an activator of Xist. As previously reported, Xist is highly expressed in the serum and monocytes of patients affected with osteoporosis and induces inhibition of the osteogenic potential of bone marrow mesenchymal stem cells, inhibiting ALP, RUNX2, and osteocalcin expression [79].
Bone-resident mast cell proliferation, activation and degranulation are involved in the pathogenesis of osteoporosis [125]. The lncRNA profile of EVs from bone marrow-derived degranulated mast cells was analyzed by Liang and colleagues [126]. Interestingly, EVs were enriched in lncRNA MALAT-1, exerting a direct effect on macrophages/osteoclasts. Functional experiments on bone marrow macrophages revealed that, once transferred into target cells, MALAT-1 repressed miR-124, inducing overexpression of mmp9, ctsk, acp5, and car2 genes and finally increasing osteoclastogenesis and bone resorption [127].
Sun and colleagues showed that osteoclasts secrete miR-214-enriched exosomes [128]. These exosomes are recognized via the EphrinA2/EphA2 axis and integrated by osteoblasts, inhibiting their activity. Exosomes from both OVX mice and osteoporotic patients contained high levels of miR-214 and ephrinA2 protein, suggesting that osteoclastic exosomes exert a noticeable inhibitory function on osteoblast activity in vivo. This mechanism was confirmed in OVX mice, since the prevention of exosome formation by downregulation of Rab27a increased osteoblast activity.
5.3. Endothelial-Derived EVs
Angiogenesis and bone remodeling are tightly related processes [129,130]. Both these processes can be heavily affected by senescence; in fact, both vascular and bone disfunction, as well as many other syndromes, can be alleviated by senolytic agents [131,132,133,134,135,136]. Weilner et al. demonstrated that miR-31 is present at elevated levels in the plasma of an elderly population [137]. Similarly, they reported serum miR-31 is elevated in the plasma of osteoporosis patients, therefore hypothesizing that miR-31 in the plasma of the elderly might play a role in the pathogenesis of age-related impaired bone formation. Looking at the source of miR-31, Weilner et al. identified senescent endothelial cells as producers, delivering miR-31 within EVs [137]. When miR-31 is taken up by mesenchymal stem cells, it inhibits osteogenic differentiation by counteracting its target Frizzled-3 gene.
Endothelial cells can also affect osteoclastogenesis via EVs [127]. Endothelial progenitor cells secrete sEVs able to promote recruitment and differentiation of osteoclast precursors. These sEVs are enriched in lncRNA MALAT 1, exhibiting a pro-osteoclastogenic effect, as already discussed.
5.4. Muscle-Derived EVs
Bone homeostasis is tightly correlated with muscle performance and mass [138,139,140]. Along with the mechanic stimulation on the bone due to the kinetics of muscle contraction, muscle cells release specific cytokines, known as myokines, with anabolic/catabolic action on bone [141]. On this basis, osteosarcopenia, a term coined by Duque and colleagues, is a new emerging musculoskeletal syndrome [142]. In this context, EVs isolated from serum samples of elderly subjects showed a significant increase in a muscle-derived alpha-sarcoglycan positive EV subpopulation, compared to young controls [143]. Fulzele and colleagues showed that these EVs were also enriched in miR-34a. When they overexpressed miR-34a in mouse myoblast C2C12 cells by lentiviral vector infection and induced miR-34a human primary myotubes by hydrogen peroxide treatment (to simulate oxidative stress), the cells released EVs enriched in miR-34a, which in turn were able to abrogate Sirtuin 1 (sirt1). Sirt1 was reported also by other authors to be related to senescence in bone marrow stromal cells, decreasing osteogenic potential and bone deposition [144]. The authors concluded that their findings suggest that aged skeletal muscle is a potential source of circulating, senescence-associated EVs that may directly impact stem cell populations in tissues such as bone via their microRNA cargo.
Serum levels of muscle-derived cytokine myostatin increase with progressing age both in men and in women and are inversely associated with skeletal muscle mass [145,146]. Qin et al. showed that high levels of myostatin affect bone cells, especially osteocytes. In particular, Qui et al. described that the osteocytic cells Ocy454 and their sEVs contain miR-218 in basal condition [147]. Upon exposition of the myostatin, the Ocy454 cells increase SOST, DKK1, and RANKL production and decrease miR-218 expression. Interestingly, consequently to the cytoplasmatic drop in miR-218, the miR-218 content decreased in their sEVs. In addition, the same authors showed that osteoblast-like MC3T3 cells treated with miR-218-depleted osteocyte sEVs exhibited a stunted osteogenic differentiation. Finally, to confirm these data, Qin et al. demonstrated that exogenous overexpression of miR-218 in MC3T3 cells rescued the effect of myostatin-treated osteocyte sEVs, avoiding the increase in SOST and RANKL production, and the suppression of osteoblast differentiation and function. The authors concluded that, in addition to the effect on bone remodeling exerted through SOST and RANKL, the osteocytes can contribute to dampening osteogenesis by sEVs depleted in miRNA-218, adding one more EV-based mechanism contributing to altering the molecular asset of osteoblast function and osteogenesis.
A summary of all the miRNAs described above and their effects is reported in Table 1.
6. The Other Side of the Coin: EVs as a Potential Clinical Tool for Osteoporosis
In parallel with the studies elucidating the negative effect of EVs on bone metabolism and their involvement in the progression of skeletal diseases such as osteoporosis, many other reports highlighted a positive role of some EVs on bone metabolism, paving the way for their potential use in bone tissue engineering. This discrepancy can be explained by the difference in the molecular content of EVs, mirroring the status of the donor cells, as well as the asset of the target cells. Cui et al. showed that sEVs derived from mineralizing pre-osteoblast MC3T3-E1 cells can promote bone marrow stromal cell (ST2 cell) differentiation towards osteoblast [148]. Once taken up by ST2 cells, EVs induce a change in the miRNA profile of ST2, abrogating Axin1 expression while increasing β-catenin expression, overall leading to the activation of Wnt signaling and osteogenesis.
A study by Guo and colleagues demonstrated that sEVs from human synovial-derived mesenchymal stem cells (SMSCs) alleviate glucocorticoid-induced osteonecrosis of the femoral head (ONFH) in an experimental rat model [149]. Upon in vivo infusion, SMSC-EVs exert proliferative and antiapoptotic effects on bone marrow cells and improve bone mineral density and microarchitecture of femurs, increasing the number of osteogenic cells.
More recently, Liao and colleagues dissected the molecular mechanism of the ONFH progression in a rabbit model and the potential therapeutic use of mesenchymal stromal cell-derived EVs [150]. The authors showed that miR-122 is the most downregulated miRNA in ONFH. Interestingly, the SPRY2 gene is a target for miR-122 and inhibits osteoblast differentiation via the RTK/Ras/MAPK signaling cascade suppression, and it has been reported to increase ONFH. Finally, EVs derived from bone marrow stromal cells overexpressing miR-122 were shown to attenuate ONFH development in vivo.
Endothelial cell-derived EVs were used as a preventive therapeutic tool in an experimental mouse model of steroid-induced osteoporosis [151]. The authors showed that dexamethasone induces ferroptosis in osteoblasts, suppressing glutathione peroxidase 4 (GPX4), system Xc−, and cysteine levels while upregulating malondialdehyde (MDA) and reactive oxygen species (ROS) production. EVs isolated by bone marrow-derived endothelial progenitor cells (BM-EPCs) injected in the tail vein of mice were able to mitigate the ferroptotic progression and increase bone parameters, such as bone volume, trabecular thickness, and structure model index. Although the authors did not investigate in detail the molecular players shuttled by EV-BM-EPCs, they attribute this feature to the miRNA and lncRNA content of EVs, since miR-17-92, miR-9, and miR-137, as well as lncRNA LINC00336, exhibit anti-ferroptotic activity.
7. Conclusions and Outlook
Knowledge about the function and the role of EV-based intercellular crosstalk adds one more piece in the intricate and crowded mechanism of the fine-tuned regulation of bone metabolism. It also contributes to understanding the molecular basis of the onset and progression of bone diseases. Indeed, EVs are an exciting tool with brilliant perspectives for clinical and theranostic development. These tiny structures and their molecular content could be used as biomarkers for diagnosis and monitoring of osteoporosis, as well as predictive and prognostic signatures.
Currently, some important limitations still dampen their use beyond the basic research. Major technical hurdles are still limiting the use of EVs as diagnostic tools [152]. The identification of the proper or best matrix or source (serum, plasma, urine) and the collection and the manipulation of the sample are still an unsolved aspect [153,154,155]. Furthermore, both the EVs and their molecular content can be modified by several patient-related variables, including the circadian rhythm and lifestyle habits such as physical activity [156,157,158]. Another important source of bias for EV-related RNA evaluation stems from the analytical platform, since results obtained from different technologies reported significant discrepancies in evaluating RNA levels, in conjunction with the applied normalization strategy [159,160].
However, mounting evidence supports the development of EVs as a diagnostic tool. EVs offer the invaluable possibility to obtain tissue-specific information with a non-invasive sampling procedure, the so-called liquid biopsy [161]. The stability of the EV molecular content is remarkable, since it can remain unaltered for many years when stored at controlled temperatures and is unchanged at extreme pH values and after several cycles of thawing/freezing [162,163]. The potential utility of EVs as a biomarker for bone diseases is further highlighted by the limitations of currently available diagnostic tools. The typical bone turnover markers (C-terminal type I collagen crosslinks (CTX), type I pro-collagen pro-peptide (PINP1), pyridinoline/deoxypyridinoline, parathyroid hormone (PTH), osteocalcin, bone alkaline phosphatase (BALP), and tartrate-resistant acid phosphatase 5b (TRAP5b)) are not completely accurate in evaluating bone formation and reabsorption, and dual-energy X-ray absorptiometry (DXA) provides information only about bone quality once important architectural alterations have occurred, often being irreversible [163]. Thus, we lack really efficient diagnostic tools to predict the risk or the evolution of metabolic bone diseases as well as the response to the treatments. Circulating EVs are promising biomarkers since they inform about epigenetic and transcriptomic modifications, making it possible to anticipate the alterations in the downstream signaling cascades and in classical protein markers.
EVs has been also proposed as a pharmacological platform for the treatment of musculoskeletal diseases [164]. EVs present attractive therapeutic potentiality due to a long half-life, high biocompatibility, and minimal or no adverse effects. Moreover, native (or engineered) EVs can be administrated to transport endogenous (or exogenous) molecules by their ability to target specific tissues. Regenerative medicine and targeted delivery of drugs benefit the most from the versatility of EVs. Degenerative and inflammatory disease, such as osteoporosis, can benefit from the therapeutic potential of EVs. EVs from mouse vascular endothelial cells are enriched in miR-155, exhibiting protective effects on bone in an ovariectomized (OVX) mice model of menopausal osteoporosis [165]. The pro-osteogenic potential of EVs released by mouse MSCs was specifically addressed to the bone by linking the murine MSC-derived EVs with alendronate [166]. These bone-targeting EVs were effective in protecting bone of OVX mice. In another study, murine osteoblast EVs were loaded with zoledronic acid, being able to suppress exacerbated bone resorption in mice [106]. However, some concerns (as reported for the diagnostic approaches) still limit EVs to becoming a concrete therapeutic system.
Author Contributions
Conceptualization, A.C.; original draft, A.C.; review and editing, M.M. and A.C.; funding acquisition, M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This paper and the APC were funded by Ministero della Salute: “Ricerca Finalizzata #RF-20130235739 to M.M.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Compston, J.E.; McClung, M.R.; Leslie, W.D. Osteoporosis. Lancet 2019, 393, 364–376. [Google Scholar] [CrossRef]
- Genant, H.K.; Cooper, C.; Poor, G.; Reid, I.; Ehrlich, G.; Kanis, J.; Nordin, B.E.C.; Barrett-Connor, E.; Black, D.; Bonjour, J.P.; et al. Interim report and recommendations of the World Health Organization Task-Force for Osteoporosis. Osteoporos. Int. 1999, 10, 259–264. [Google Scholar] [CrossRef]
- Al Anouti, F.; Taha, Z.; Shamim, S.; Khalaf, K.; Al Kaabi, L.; Alsafar, H. An insight into the paradigms of osteoporosis: From genetics to biomechanics. Bone Rep. 2019, 11, 100216. [Google Scholar] [CrossRef] [PubMed]
- Pisani, P.; Renna, M.D.; Conversano, F.; Casciaro, E.; Di Paola, M.; Quarta, E.; Muratore, M.; Casciaro, S. Major osteoporotic fragility fractures: Risk factor updates and societal impact. World J. Orthop. 2016, 7, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Zebaze, R.M.; Ghasem-Zadeh, A.; Bohte, A.; Iuliano-Burns, S.; Mirams, M.; Price, R.I.; Mackie, E.J.; Seeman, E. Intracortical remodelling and porosity in the distal radius and post-mortem femurs of women: A cross-sectional study. Lancet 2010, 375, 1729–1736. [Google Scholar] [CrossRef]
- Langdahl, B.; Ferrari, S.; Dempster, D.W. Bone modeling and remodeling: Potential as therapeutic targets for the treatment of osteoporosis. Ther. Adv. Musculoskelet. Dis. 2016, 8, 225–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eriksen, E.F.; Hodgson, S.F.; Eastell, R.; Riggs, B.L.; Cedel, S.L.; O’Fallon, W.M. Cancellous bone remodeling in type i (postmenopausal) osteoporosis: Quantitative assessment of rates of formation, resorption, and bone loss at tissue and cellular levels. J. Bone Miner. Res. 1990, 5, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Adler, R.A. Osteoporosis in men: A review. Bone Research 2014, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Richards, J.B.; Zheng, H.F.; Spector, T.D. Genetics of osteoporosis from genome-wide association studies: Advances and challenges. Nat. Rev. Genet. 2012, 13, 576–588. [Google Scholar] [CrossRef]
- Wu, Q.-Y.; Li, X.; Miao, Z.-N.; Ye, J.-X.; Wang, B.; Zhang, F.; Xu, R.-S.; Jiang, D.-L.; Zhao, M.-D.; Yuan, F.L. Long Non-coding RNAs: A New Regulatory Code for Osteoporosis. Front. Endocrinol. 2018, 9, 587. [Google Scholar] [CrossRef] [Green Version]
- Bellavia, D.; De Luca, A.; Carina, V.; Costa, V.; Raimondi, L.; Salamanna, F.; Alessandro, R.; Fini, M.; Giavaresi, G. Deregulated miRNAs in bone health: Epigenetic roles in osteoporosis. Bone 2019, 122, 52–75. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Wu, K.-H.; Liu, K.; Hu, Y.; Chen, X.-D.; Zhang, L.H.; Shen, H.; Tian, Q.; Zhao, L.-J.; Deng, H.-W.; et al. Genome-wide association study of lncRNA polymorphisms with bone mineral density. Ann. Hum. Genet. 2018, 82, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liang, H.; Kourkoumelis, N.; Wu, Z.; Li, G.; Shang, X. Comprehensive Analysis of lncRNA and miRNA Expression Profiles and ceRNA Network Construction in Osteoporosis. Calcif. Tissue Int. 2020, 106, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Cappariello, A.; Ponzetti, M.; Rucci, N. The ‘soft’ side of the bone: Unveiling its endocrine functions. Horm. Mol. Biol. Clin. Investig. 2016, 28, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Cappariello, A.; Maurizi, A.; Veeriah, V.; Teti, A. Reprint of: The Great Beauty of the osteoclast. Arch. Biochem. Biophys. 2014, 561, 13–21. [Google Scholar] [CrossRef]
- Parfitt, A.M. Osteonal and hemiosteonal remodeling: The spatial and temporal framework for signal traffic in adult human bone. J. Cell. Biochem 1994, 55, 273–286. [Google Scholar] [CrossRef]
- Sims, N.A.; Martin, T.J. Coupling the activities of bone formation and resorption: A multitude of signals within the basic multicellular unit. Bonekey Rep. 2014, 3, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rowe, P.; Sharma, S. Physiology, Bone Remodeling; StatPearls Publishing: Treasure Island, FL, USA, 2018. [Google Scholar]
- Robling, A.G.; Bonewald, L.F. The Osteocyte: New Insights. Annu. Rev. Physiol. 2020, 82, 485–506. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, Z.; Fu, Q.; Zhang, J. Plasma miRNA levels correlate with sensitivity to bone mineral density in postmenopausal osteoporosis patients. Biomarkers 2014, 19, 553–556. [Google Scholar] [CrossRef]
- Cappariello, A.; Berardi, A.C.; Peruzzi, B.; Del Fattore, A.; Ugazio, A.; Bottazzo, G.F.; Teti, A. Committed osteoclast precursors colonize the bone and improve the phenotype of a mouse model of autosomal recessive osteopetrosis. J. Bone Miner. Res. 2010, 25, 106–113. [Google Scholar] [CrossRef]
- Intemann, J.; De Gorter, D.J.J.; Naylor, A.J.; Dankbar, B.; Wehmeyer, C. Importance of osteocyte-mediated regulation of bone remodelling in inflammatory bone disease. Swiss Med. Wkly. 2020, 150, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.; Laurent, M.R.; Dubois, V.; Claessens, F.; O’Brien, C.A.; Bouillon, R.; Vanderschueren, D.; Manolagas, S.C. Estrogens and Androgens in Skeletal Physiology and Pathophysiology. Physiol. Rev. 2017, 97, 135–187. [Google Scholar] [CrossRef] [PubMed]
- Carson, J.A.; Manolagas, S.C. Effects of sex steroids on bones and muscles: Similarities, parallels, and putative interactions in health and disease. Bone 2015, 80, 67–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.-M.; Woo, J.; Wu, S.-H.; Ho, S.C. The role of vitamin D in blood pressure, endothelial and renal function in postmenopausal women. Nutrients 2013, 5, 2590–2610. [Google Scholar] [CrossRef] [PubMed]
- Duque, G.; El Abdaimi, K.; Macoritto, M.; Miller, M.M.; Kremer, R. Estrogens (E2) regulate expression and response of 1,25-dihydroxyvitamin D3 receptors in bone cells: Changes with aging and hormone deprivation. Biochem. Biophys. Res. Commun. 2002, 299, 446–454. [Google Scholar] [CrossRef]
- He, H.; Liu, Y.; Tian, Q.; Papasian, C.J.; Hu, T.; Deng, H.W. Relationship of sarcopenia and body composition with osteoporosis. Osteoporos. Int. 2016, 27, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Carina, V.; Della Bella, E.; Costa, V.; Bellavia, D.; Veronesi, F.; Cepollaro, S.; Fini, M.; Giavaresi, G. Bone’s Response to Mechanical Loading in Aging and Osteoporosis: Molecular Mechanisms. Calcif. Tissue Int. 2020, 107, 301–318. [Google Scholar] [CrossRef]
- Severinsen, M.C.K.; Pedersen, B.K. Muscle-Organ Crosstalk: The Emerging Roles of Myokines. Endocr. Rev. 2020, 41, bnaa016. [Google Scholar] [CrossRef]
- McClung, M.; Harris, S.T.; Miller, P.D.; Bauer, D.C.; Davison, K.S.; Dian, L.; Hanley, D.A.; Kendler, D.L.; Yuen, C.K.; Lewiecki, E.M. Bisphosphonate therapy for osteoporosis: Benefits, risks, and drug holiday. Am. J. Med. 2013, 126, 13–20. [Google Scholar] [CrossRef]
- Cummings, S.R.; San Martin, J.; McClung, M.R.; Siris, E.S.; Eastell, R.; Reid, I.R.; Delmas, P.; Zoog, H.B.; Austin, M.; Wang, A.; et al. Denosumab for Prevention of Fractures in Postmenopausal Women with Osteoporosis. N. Engl. J. Med. 2009, 361, 756–765. [Google Scholar] [CrossRef] [Green Version]
- Tu, K.N.; Lie, J.D.; Wan, C.K.V.; Cameron, M.; Austel, A.G.; Nguyen, J.K.; Van, K.; Hyun, D. Osteoporosis: A review of treatment options. Pharm. Ther. 2018, 43, 92–104. [Google Scholar]
- Farrah, Z.; Jawad, A.S.M. Optimising the management of osteoporosis. Clin. Med. J. R. Coll. Physicians Lond. 2020, 20, E196–E201. [Google Scholar]
- Cauley, J.A.; Robbins, J.; Chen, Z.; Cummings, S.R.; Jackson, R.D.; LaCroix, A.Z.; LeBoff, M.; Lewis, C.E.; McGowan, J.; Neuner, J.; et al. Effects of Estrogen Plus Progestin on Risk of Fracture and Bone Mineral Density: The Women’s Health Initiative Randomized Trial. J. Am. Med. Assoc. 2003, 290, 1729–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodis, H.N.; Mack, W.J.; Henderson, V.W.; Shoupe, D.; Budoff, M.J.; Hwang-Levine, J.; Li, Y.; Feng, M.; Dustin, L.; Kono, N.; et al. Vascular Effects of Early versus Late Postmenopausal Treatment with Estradiol. N. Engl. J. Med. 2016, 374, 1221–1231. [Google Scholar] [CrossRef] [PubMed]
- Riggs, B.L.; Hartmann, L.C. Selective estrogen-receptor modulators—Mechanisms of action and application to clinical practice. N. Engl. J. Med. 2003, 348, 618–629. [Google Scholar] [CrossRef]
- Cummings, S.R.; Eckert, S.; Krueger, K.A.; Grady, D.; Powles, T.J.; Cauley, J.A.; Norton, L.; Nickelsen, T.; Bjarnason, N.H.; Morrow, M.; et al. The effect of raloxifene on risk of breast cancer in postmenopausal women: Results from the MORE randomized trial. J. Am. Med. Assoc. 1999, 281, 2189–2197. [Google Scholar] [CrossRef] [Green Version]
- Miller, P.D.; Hattersley, G.; Riis, B.J.; Williams, G.C.; Lau, E.; Russo, L.A.; Alexandersen, P.; Zerbini, C.A.; Hu, M.Y.; Harris, A.G.; et al. Effect of abaloparatide vs. placebo on newvertebral fractures in postmenopausalwomen with osteoporosis a randomized clinical trial. JAMA J. Am. Med. Assoc. 2016, 316, 722–733. [Google Scholar] [CrossRef]
- Dempster, D.W.; Zhou, H.; Ruff, V.A.; Melby, T.E.; Alam, J.; Taylor, K.A. Longitudinal Effects of Teriparatide or Zoledronic Acid on Bone Modeling- and Remodeling-Based Formation in the SHOTZ Study. J. Bone Miner. Res. 2018, 33, 627–633. [Google Scholar] [CrossRef] [Green Version]
- Saag, K.G.; Petersen, J.; Brandi, M.L.; Karaplis, A.C.; Lorentzon, M.; Thomas, T.; Maddox, J.; Fan, M.; Meisner, P.D.; Grauer, A. Romosozumab or Alendronate for Fracture Prevention in Women with Osteoporosis. N. Engl. J. Med. 2017, 377, 1417–1427. [Google Scholar] [CrossRef] [Green Version]
- Bird, A. Perceptions of epigenetics. Nature 2007, 447, 396–398. [Google Scholar] [CrossRef]
- Egger, G.; Liang, G.; Aparicio, A.; Jones, P.A. Epigenetics in human disease and prospects for epigenetic therapy. Nature 2004, 429, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Peschansky, V.J.; Wahlestedt, C. Non-coding RNAs as direct and indirect modulators of epigenetic regulation. Epigenetics 2014, 9, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaratiegui, M.; Irvine, D.V.; Martienssen, R.A. Noncoding RNAs and Gene Silencing. Cell 2007, 128, 763–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and Functions of Long Noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Fabian, M.R.; Sonenberg, N. The mechanics of miRNA-mediated gene silencing: A look under the hood of miRISC. Nat. Struct. Mol. Biol. 2012, 19, 586–593. [Google Scholar] [CrossRef]
- Luteijn, M.J.; Ketting, R.F. PIWI-interacting RNAs: From generation to transgenerational epigenetics. Nat. Rev. Genet. 2013, 14, 523–534. [Google Scholar] [CrossRef] [Green Version]
- Okamura, K.; Chung, W.J.; Ruby, J.G.; Guo, H.; Bartel, D.P.; Lai, E.C. The Drosophila hairpin RNA pathway generates endogenous short interfering RNAs. Nature 2008, 453, 803–806. [Google Scholar] [CrossRef]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [Green Version]
- Treps, L.; Perret, R.; Edmond, S.; Ricard, D.; Gavard, J. Glioblastoma stem-like cells secrete the proangiogenic VEGF-A factor in extracellular vesicles. J. Extracell. Vesicles 2017, 6, 1359479. [Google Scholar] [CrossRef]
- Engreitz, J.M.; Haines, J.E.; Perez, E.M.; Munson, G.; Chen, J.; Kane, M.; McDonel, P.E.; Guttman, M.; Lander, E.S. Local regulation of gene expression by lncRNA promoters, transcription and splicing. Nature 2016, 539, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS ONE 2012, 7, e30733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.C.R.; Acuña, S.M.; Aoki, J.I.; Floeter-Winter, L.M.; Muxel, S.M. Long non-coding RNAs in the regulation of gene expression: Physiology and disease. Non Coding RNA 2019, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Amaral, P.P.; Dinger, M.E.; Mercer, T.R.; Mattick, J.S. The eukaryotic genome as an RNA machine. Science 2008, 319, 1787–1789. [Google Scholar] [CrossRef]
- Costa, F.F. Non-coding RNAs, epigenetics and complexity. Gene 2008, 410, 9–17. [Google Scholar] [CrossRef]
- Ghildiyal, M.; Zamore, P.D. Small silencing RNAs: An expanding univers. Nat. Rev. Genet. 2009, 10, 94–108. [Google Scholar] [CrossRef] [Green Version]
- Ruvkun, G. Molecular biology: Glimpses of a tiny RNA world. Science 2001, 294, 797–799. [Google Scholar] [CrossRef] [Green Version]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [Green Version]
- Griffiths-Jones, S.; Grocock, R.J.; van Dongen, S.; Bateman, A.; Enright, A.J. miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuddenham, L.; Wheeler, G.; Ntounia-Fousara, S.; Waters, J.; Hajihosseini, M.K.; Clark, I.; Dalmay, T. The cartilage specific microRNA-140 targets histone deacetylase 4 in mouse cells. FEBS Lett. 2006, 580, 4214–4217. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, M.; Garzon, R.; Cimmino, A.; Liu, Z.; Zanesi, N.; Callegari, E.; Liu, S.; Alder, H.; Costinean, S.; Fernandez-Cymering, C.; et al. MicroRNA-29 family reverts aberrant methylation in lung cancer by targeting DNA methyltransferases 3A and 3B. Proc. Natl. Acad. Sci. USA 2007, 104, 15805–15810. [Google Scholar] [CrossRef] [Green Version]
- Seeliger, C.; Karpinski, K.; Haug, A.T.; Vester, H.; Schmitt, A.; Bauer, J.S.; van Griensven, M. Five Freely Circulating miRNAs and Bone Tissue miRNAs Are Associated with Osteoporotic Fractures. J. Bone Miner. Res. 2014, 29, 1718–1728. [Google Scholar] [CrossRef]
- Weilner, S.; Skalicky, S.; Salzer, B.; Keider, V.; Wagner, M.; Hildner, F.; Gabriel, C.; Dovjak, P.; Pietschmann, P.; Grillari-Voglauer, R.; et al. Differentially circulating miRNAs after recent osteoporotic fractures can influence osteogenic differentiation. Bone 2015, 79, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Kung, J.T.Y.; Colognori, D.; Lee, J.T. Long noncoding RNAs: Past, present, and future. Genetics 2013, 193, 651–669. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.C.; Chang, H.Y. Molecular Mechanisms of Long Noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [Green Version]
- Dykes, I.M.; Emanueli, C. Transcriptional and Post-transcriptional Gene Regulation by Long Non-coding RNA. Genom. Proteom. Bioinform. 2017, 15, 177–186. [Google Scholar] [CrossRef]
- Liang, W.C.; Fu, W.M.; Wang, Y.B.; Sun, Y.X.; Xu, L.L.; Wong, C.W.; Chan, K.M.; Li, G.; Waye, M.M.; Zhang, J.F. H19 activates Wnt signaling and promotes osteoblast differentiation by functioning as a competing endogenous RNA. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef]
- Yuan, X.; Guo, Y.; Chen, D.; Luo, Y.; Chen, D.; Miao, J.; Chen, Y. Long non-coding RNA MALAT1 functions as miR-1 sponge to regulate Connexin 43 mediated ossification of the posterior longitudinal ligament. Bone 2019, 127, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Wu, J.; Zheng, Z.; Wang, G.; Song, D.; Yan, P.; Yin, H.; Hu, P.; Zhu, X.; Wang, H.; et al. The Construction and Analysis of ceRNA Network and Patterns of Immune Infiltration in Mesothelioma with Bone Metastasis. Front. Bioeng. Biotechnol. 2019, 7, 257. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.M.; Moura, S.R.; Teixeira, J.H.; Barbosa, M.A.; Santos, S.G.; Almeida, M.I. Long noncoding RNAs: A missing link in osteoporosis. Bone Res. 2019, 7, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercer, T.R.; Mattick, J.S. Structure and function of long noncoding RNAs in epigenetic regulation. Nat. Struct. Mol. Biol. 2013, 20, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Bunch, H. Gene regulation of mammalian long non-coding RNA. Mol. Genet. Genom. 2018, 293, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Maass, P.G.; Luft, F.C.; Bähring, S. Long non-coding RNA in health and disease. J. Mol. Med. 2014, 92, 337–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Yang, L.; Ge, D.; Wang, W.; Yin, Z.; Yan, J.; Cao, X.; Jiang, C.; Zheng, S.; Liang, B. Long non-coding RNA XIST promotes osteoporosis through inhibiting bone marrow mesenchymal stem cell differentiation. Exp. Ther. Med. 2019, 17, 803–811. [Google Scholar] [CrossRef]
- Mei, B.; Wang, Y.; Ye, W.; Huang, H.; Zhou, Q.; Chen, Y.; Niu, Y.; Zhang, M.; Huang, Q. LncRNA ZBTB40-IT1 modulated by osteoporosis GWAS risk SNPs suppresses osteogenesis. Hum. Genet. 2019, 138, 151–166. [Google Scholar] [CrossRef]
- Li, B.; Liu, J.; Zhao, J.; Ma, J.X.; Jia, H.B.; Zhang, Y.; Xing, G.S.; Ma, X.L. LncRNA-H19 Modulates Wnt/β-catenin Signaling by Targeting Dkk4 in Hindlimb Unloaded Rat. Orthop. Surg. 2017, 9, 319–327. [Google Scholar] [CrossRef]
- Wolf, P. The nature and significance of platelet products in human plasma. Br. J. Haematol. 1967, 13, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.T.; Teng, K.; Wu, C.; Adam, M.; Johnstone, R.M. Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes. J. Cell Biol. 1985, 101, 942–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashed, M.H.; Bayraktar, E.; Helal, G.K.; Abd-Ellah, M.F.; Amero, P.; Chavez-Reyes, A.; Rodriguez-Aguayo, C. Exosomes: From garbage bins to promising therapeutic targets. Int. J. Mol. Sci. 2017, 18, 538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, Secretion, and Intercellular Interactions of Exosomes and Other Extracellular Vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef] [PubMed]
- Poon, I.K.H.; Hulett, M.D.; Parish, C.R. Molecular mechanisms of late apoptotic/necrotic cell clearance. Cell Death Differ. 2010, 17, 381–397. [Google Scholar] [CrossRef] [Green Version]
- Atkin-Smith, G.K.; Tixeira, R.; Paone, S.; Mathivanan, S.; Collins, C.; Liem, M.; Goodall, K.J.; Ravichandran, K.S.; Hulett, M.D.; Poon, I.K. A novel mechanism of generating extracellular vesicles during apoptosis via a beads-on-a-string membrane structure. Nat. Commun. 2015, 6, 1–10. [Google Scholar] [CrossRef]
- Cappariello, A.; Loftus, A.; Muraca, M.; Maurizi, A.; Rucci, N.; Teti, A. Osteoblast-Derived ExtracellularVesicles Are Biological Tools for the Delivery of Active Molecules to Bone. J. Bone Miner. Res. 2018, 33, 517–533. [Google Scholar] [CrossRef] [Green Version]
- Pieters, B.C.H.; Cappariello, A.; van den Bosch, M.H.J.; van Lent, P.L.E.M.; Teti, A.; van de Loo, F.A.J. Macrophage-derived extracellular vesicles as carriers of alarmins and their potential involvement in bone homeostasis. Front. Immunol. 2019, 10, 1901. [Google Scholar] [CrossRef] [Green Version]
- Ikebuchi, Y.; Aoki, S.; Honma, M.; Hayashi, M.; Sugamori, Y.; Khan, M.; Kariya, Y.; Kato, G.; Tabata, Y.; Penninger, J.M.; et al. Coupling of bone resorption and formation by RANKL reverse signalling. Nature 2018, 561, 195–200. [Google Scholar] [CrossRef]
- Sato, M.; Suzuki, T.; Kawano, M.; Tamura, M. Circulating osteocyte-derived exosomes contain miRNAs which are enriched in exosomes from MLO-Y4 cells. Biomed. Rep. 2017, 6, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Roig-Arcos, J.; López-Malo, D.; Díaz-Llopis, M.; Romero, F.J. Exosomes derived from stimulated monocytes promote endothelial dysfunction and inflammation in vitro. Ann. Transl. Med. 2017, 5, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, M.N.A.; Nieuwland, R.; Hau, C.M.; Evers, L.M.; Meesters, E.W.; Sturk, A. Cell-derived microparticles contain caspase 3 in vitro and in vivo. J. Thromb. Haemost. 2005, 3, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Loftus, A.; Cappariello, A.; George, C.; Ucci, A.; Shefferd, K.; Green, A.; Paone, R.; Ponzetti, M.; Delle Monache, S.; Muraca, M.; et al. Extracellular Vesicles from Osteotropic Breast Cancer Cells Affect Bone Resident Cells. J. Bone Miner. Res. 2020, 35, 396–412. [Google Scholar] [CrossRef] [Green Version]
- Cappariello, A.; Rucci, N. Tumour-derived extracellular vesicles (EVs): A dangerous ‘message in a bottle’ for bone. Int. J. Mol. Sci. 2019, 20, 4805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yáñez-Mó, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 1–60. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, D.; Chen, X.; Li, J.; Li, L.; Bian, Z.; Sun, F.; Lu, J.; Yin, Y.; Cai, X.; et al. Secreted Monocytic miR-150 Enhances Targeted Endothelial Cell Migration. Mol. Cell 2010, 39, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Lv, L.L.; Feng, Y.; Wu, M.; Wang, B.; Li, Z.L.; Zhong, X.; Wu, W.J.; Chen, J.; Ni, H.F.; Tang, T.T.; et al. Exosomal miRNA-19b-3p of tubular epithelial cells promotes M1 macrophage activation in kidney injury. Cell Death Differ. 2020, 27, 210–226. [Google Scholar] [CrossRef]
- Umezu, T.; Tadokoro, H.; Azuma, K.; Yoshizawa, S.; Ohyashiki, K.; Ohyashiki, J.H. Exosomal miR-135b shed from hypoxic multiple myeloma cells enhances angiogenesis by targeting factor-inhibiting HIF-1. Blood 2014, 124, 3748–3757. [Google Scholar] [CrossRef]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef]
- Anderson, H.C. Electron microscopic studies of induced cartilage development and calcification. J. Cell Biol. 1967, 35, 81–101. [Google Scholar] [CrossRef] [PubMed]
- Bonucci, E. Fine structure of early cartilage calcification. J. Ultrasructure Res. 1967, 20, 33–50. [Google Scholar] [CrossRef]
- Davies, O.G.; Cox, S.C.; Williams, R.L.; Tsaroucha, D.; Dorrepaal, R.M.; Lewis, M.P.; Grover, L.M. Annexinenriched osteoblast-derived vesicles act as an extracellular site of mineral nucleation within developing stem cell cultures. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.Y.; Yoshida, H.; Sarosi, I.; Tan, H.L.; Timms, E.; Capparelli, C.; Morony, S.; Oliveira-dos-Santos, A.J.; Van, G.; Itie, A.; et al. OPGL is a key regulator of osteoclastogenesis, lymphocyte development and lymph-node organogenesis. Nature 1999, 397, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Cappariello, A.; Paone, R.; Maurizi, A.; Capulli, M.; Rucci, N.; Muraca, M.; Teti, A. Biotechnological approach for systemic delivery of membrane Receptor activator of NF-κB Ligand (RANKL) active domain into the circulation. Biomaterials 2015, 46, 58–69. [Google Scholar] [CrossRef]
- Del Fattore, A.; Cappariello, A.; Capulli, M.; Rucci, N.; Muraca, M.; De Benedetti, F.; Teti, A. An experimental therapy to improve skeletal growth and prevent bone loss in a mouse model overexpressing IL-6. Osteoporos. Int. 2014, 25, 681–692. [Google Scholar] [CrossRef]
- Deng, L.; Wang, Y.; Peng, Y.; Wu, Y.; Ding, Y.; Jiang, Y.; Shen, Z.; Fu, Q. Osteoblast-derived microvesicles: A novel mechanism for communication between osteoblasts and osteoclasts. Bone 2015, 79, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Weilner, S.; Keider, V.; Winter, M.; Harreither, E.; Salzer, B.; Weiss, F.; Schraml, E.; Messner, P.; Pietschmann, P.; Hildner, F.; et al. Vesicular Galectin-3 levels decrease with donor age and contribute to the reduced osteo-inductive potential of human plasma derived extracellular vesicles. Aging 2016, 1, 16–33. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, R.; Tanaka, C.; Sato, M.; Nagasaki, H.; Sugimura, K.; Okumura, K.; Nakagawa, Y.; Aoki, N. Adipocyte-derived microvesicles contain RNA that is transported into macrophages and might be secreted into blood circulation. Biochem. Biophys. Res. Commun. 2010, 398, 723–729. [Google Scholar] [CrossRef]
- Aoki, N.; Rumi, Y.; Noriyuki, A.; Makiko, O.; Yuichi, O.; Kaori, K.; Beate, H.; Koichi, H.; Yoshimi, N.; Tsukasa, M. Adipocyte-Derived Microvesicles Are Associated with Multiple Angiogenic Factors and Induce Angiogenesis in Vivo and in Vitro. Endocrinology 2010, 151, 2567–2576. [Google Scholar] [CrossRef] [Green Version]
- Mack, M.; Kleinschmidt, A.; Brühl, H.; Klier, C.; Nelson, P.J.; Cihak, J.; Plachý, J.; Stangassinger, M.; Erfle, V.; Schlöndorff, D. Transfer of the chemokine receptor CCR5 between cells by membrane—Derived microparticles: A mechanism for cellular human immunodeficiency virus 1 infection. Nat. Med. 2000, 6, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Yang, J.; Zheng, Y.; Chen, X.; Fang, S. Extracellular vesicles from endothelial progenitor cells prevent steroid-induced osteoporosis by suppressing the ferroptotic pathway in mouse osteoblasts based on bioinformatics evidence. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef]
- Xu, J.-F.; Yang, G.H.; Pan, X.H.; Zhang, S.J.; Zhao, C.; Qiu, B.S.; Gu, H.F.; Hong, J.F.; Cao, L.; Chen, Y.; et al. Altered MicroRNA Expression Profile in Exosomes during Osteogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells. PLoS ONE 2014, 9, e114627. [Google Scholar] [CrossRef]
- Xie, Y.; Chen, Y.; Zhang, L.; Ge, W.; Tang, P. The roles of bone-derived exosomes and exosomal microRNAs in regulating bone remodelling. J. Cell Mol. Med. 2017, 21, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hassan, M.Q.; Volinia, S.; van Wijnen, A.J.; Stein, J.L.; Croce, C.M.; Lian, J.B.; Stein, G.S. A microRNA signature for a BMP2-induced osteoblast lineage commitment program. Proc. Natl. Acad. Sci. USA 2008, 105, 13906–13911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Huang, M.; Zhang, X.; Xu, J.; Hu, G.; Zhao, X.; Cui, P.; Zhang, X. MiR-146a Deletion Protects from Bone Loss in OVX Mice by Suppressing RANKL/OPG and M-CSF in Bone Microenvironment. J. Bone Miner. Res. 2019, 34, 2149–2161. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Meng, X.; Han, X.; Li, J. Exosomes derived from circRNA Rtn4-modified BMSCs attenuate TNF-αinduced cytotoxicity and apoptosis in murine MC3T3-E1 cells by sponging miR-146a. Biosci. Rep. 2020, 40, 3436. [Google Scholar] [CrossRef]
- Alexander, M.; Hu, R.; Runtsch, M.C.; Kagele, D.A.; Mosbruger, T.L.; Tolmachova, T.; Seabra, M.C.; Round, J.L.; Ward, D.M.; O’Connell, R.M. Exosome-delivered microRNAs modulate the inflammatory response to endotoxin. Nat. Commun. 2015, 6, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Zhou, Q.; Fu, T.; Zhao, R.; Yang, J.; Kong, X.; Zhang, Z.; Sun, C.; Bao, Y.; Ge, X.; et al. Circulating Exosomes Derived-miR-146a from Systemic Lupus Erythematosus Patients Regulates Senescence of Mesenchymal Stem Cells. Biomed. Res. Int. 2019, 6071308. [Google Scholar] [CrossRef] [Green Version]
- Scheideler, M.; Elabd, C.; Zaragosi, L.E.; Chiellini, C.; Hackl, H.; Sanchez-Cabo, F.; Yadav, S.; Duszka, K.; Friedl, G.; Papak, C. Comparative transcriptomics of human multipotent stem cells during adipogenesis and osteoblastogenesis. BMC Genom. 2008, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yu, M.; Dai, M.; Chen, C.; Tang, Q.; Jing, W.; Wang, H.; Tian, W. miR-450a-5p within rat adipose tissue exosome-like vesicles promotes adipogenic differentiation by targeting WISP2. J. Cell Sci. 2017, 130, 1158–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, P.J.; Haren, N.; Ghali, O.; Clabaut, A.; Chauveau, C.; Hardouin, P.; Broux, O. Adipogenic RNAs are transferred in osteoblasts via bone marrow adipocytes-derived extracellular vesicles (EVs). BMC Cell Biol. 2015, 16, 12860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Yuan, T.; Yang, C.; Wang, Z.; Zang, Y.; Wu, L.; Zhuang, L. X-inactive-specific transcript of peripheral blood cells is regulated by exosomal Jpx and acts as a biomarker for female patients with hepatocellular carcinoma. Ther. Adv. Med. Oncol. 2017, 9, 665–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ragipoglu, D.; Dudeck, A.; Haffner-Luntzer, M.; Voss, M.; Kroner, J.; Ignatius, A.; Fischer, V. The Role of Mast Cells in Bone Metabolism and Bone Disorders. Front. Immunol. 2020, 11, 163. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Huang, S.; Qiao, L.; Peng, X.; Li, C.; Lin, K.; Xie, G.; Li, J.; Lin, L.; Yin, Y.; et al. Characterization of protein, long noncoding RNA and microRNA signatures in extracellular vesicles derived from resting and degranulated mast cells. J. Extracell. Vesicles 2020, 9, 1697583. [Google Scholar] [CrossRef]
- Cui, Y.; Fu, S.; Sun, D.; Xing, J.; Hou, T.; Wu, X. EPC-derived exosomes promote osteoclastogenesis through LncRNA-MALAT1. J. Cell. Mol. Med. 2019, 23, 3843–3854. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Zhao, C.; Li, Y.; Wang, L.; Nie, G.; Peng, J.; Wang, A.; Zhang, P.; Tian, W.; Li, Q.; et al. Osteoclast-derived microRNA-containing exosomes selectively inhibit osteoblast activity. Cell Discov. 2016, 2, 1–23. [Google Scholar] [CrossRef]
- Kusumbe, A.P.; Ramasamy, S.K.; Adams, R.H. Coupling of angiogenesis and osteogenesis by a specific vessel subtype in bone. Nature 2014, 507, 323–328. [Google Scholar] [CrossRef]
- Ramasamy, S.K.; Kusumbe, A.P.; Wang, L.; Adams, R.H. Endothelial Notch activity promotes angiogenesis and osteogenesis in bone. Nature 2014, 507, 376–380. [Google Scholar] [CrossRef] [Green Version]
- Erusalimsky, J.D.; Skene, C. Mechanisms of endothelial senescence. Exp. Physiol. 2009, 94, 299–304. [Google Scholar] [CrossRef]
- Jia, G.; Aroor, A.R.; Jia, C.; Sowers, J.R. Endothelial cell senescence in aging-related vascular dysfunction. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1802–1809. [Google Scholar] [CrossRef] [PubMed]
- Farr, J.N.; Xu, M.; Weivoda, M.M.; Monroe, D.G.; Fraser, D.G.; Onken, J.L.; Negley, B.A.; Sfeir, J.G.; Ogrodnik, M.B.; Hachfeld, C.M.; et al. Targeting cellular senescence prevents age-related bone loss in mice. Nat. Med. 2017, 23, 1072–1079. [Google Scholar] [CrossRef] [PubMed]
- Khosla, S.; Farr, J.N.; Kirkland, J.L. Inhibiting Cellular Senescence: A New Therapeutic Paradigm for AgeRelated Osteoporosis. J. Clin. Endocrinol. Metab. 2018, 103, 1282–1290. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Zhao, Q.; Zou, M.H. Targeting senescent cells to attenuate cardiovascular disease progression. Ageing Res. Rev. 2020, 60, 101072. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, J.L.; Tchkonia, T. Senolytic drugs: From discovery to translation. J. Intern. Med. 2020, 288, 518–536. [Google Scholar] [CrossRef] [PubMed]
- Weilner, S.; Schraml, E.; Wieser, M.; Messner, P.; Schneider, K.; Wassermann, K.; Micutkova, L.; Fortschegger, K.; Maier, A.B.; Westendorp, R.; et al. Secreted microvesicular miR-31 inhibits osteogenic differentiation of mesenchymal stem cells. Aging Cell 2016, 15, 744–754. [Google Scholar] [CrossRef] [Green Version]
- Kemmler, W.; Shojaa, M.; Kohl, M.; von Stengel, S. Effects of Different Types of Exercise on Bone Mineral Density in Postmenopausal Women: A Systematic Review and Meta-analysis. Calcif. Tissue Int. 2020, 107, 1–31. [Google Scholar] [CrossRef]
- Karlsson, M.; Lahti, A.; Cronholm, F.; Dencker, M.; Karlsson, C.; Rosengren, B. Daily school physical activity increases bone mass and gradually reduce the fracture risk. Lakartidningen 2019, 116, FHP7. [Google Scholar]
- Karlsson, M.K.; Rosengren, B.E. Exercise and Peak Bone Mass. Curr. Osteoporos. Rep. 2020, 18, 285–290. [Google Scholar] [CrossRef] [Green Version]
- Guo, B.; Zhang, Z.K.; Liang, C.; Li, J.; Liu, J.; Lu, A.; Zhang, B.T.; Zhang, G. Molecular Communication from Skeletal Muscle to Bone: A Review for Muscle-Derived Myokines Regulating Bone Metabolism. Calcif. Tissue Int. 2017, 100, 184–192. [Google Scholar] [CrossRef]
- Kirk, B.; Feehan, J.; Lombardi, G.; Duque, G. Muscle, Bone, and Fat Crosstalk: The Biological Role of Myokines, Osteokines, and Adipokines. Curr. Osteoporos. Rep. 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Fulzele, S.; Mendhe, B.; Khayrullin, A.; Johnson, M.; Kaiser, H.; Liu, Y.; Isales, C.M.; Hamrick, M.W.S. Muscle-derived miR-34a increases with age in circulating extracellular vesicles and induces senescence of bone marrow stem cells. Aging (Albany NY) 2019, 11, 1791–1803. [Google Scholar] [CrossRef] [PubMed]
- Zainabadi, K.; Liu, C.J.; Guarente, L. SIRT1 is a positive regulator of the master osteoblast transcription factor, RUNX2. PLoS ONE 2017, 12, e0178520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarasheski, K.E.; Bhasin, S.; Sinha-Hikim, I.; Pak-Loduca, J.; Gonzalez-Cadavid, N.F. Serum myostatinimmunoreactive protein is increased in 60-92 year old women and men with muscle wasting. J. Nutr. Health Aging 2002, 6, 343–348. [Google Scholar]
- White, T.A.; LeBrasseur, N.K. Myostatin and Sarcopenia: Opportunities and Challenges—A Mini-Review. Gerontology 2014, 60, 289–293. [Google Scholar] [CrossRef]
- Qin, Y.; Peng, Y.; Zhao, W.; Pan, J.; Ksiezak-Reding, H.; Cardozo, C.; Wu, Y.; Divieti Pajevic, P.; Bonewald, L.F.; Bauman, W.A.; et al. Myostatin inhibits osteoblastic differentiation by suppressing osteocyte-derived exosomal microRNA-218: A novel mechanism in muscle-bone communication. J. Biol. Chem. 2017, 292, 11021–11033. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Luan, J.; Li, H.; Zhou, X.; Han, J. Exosomes derived from mineralizing osteoblasts promote ST2 cell osteogenic differentiation by alteration of microRNA expression. FEBS Lett. 2016, 590, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.C.; Tao, S.C.; Yin, W.J.; Qi, X.; Sheng, J.G.; Zhang, C.Q. Exosomes from human synovial-derived mesenchymal stem cells prevent glucocorticoid-induced osteonecrosis of the femoral head in the rat. Int. J. Biol. Sci. 2016, 12, 1262–1272. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.; Ning, Y.; Xu, H.J.; Zou, W.Z.; Hu, J.; Liu, X.Z.; Yang, Y.; Li, Z.H. BMSC-derived exosomes carrying microRNA-122-5p promote proliferation of osteoblasts in osteonecrosis of the femoral head. Clin. Sci. 2019, 133, 1955–1975. [Google Scholar] [CrossRef]
- Yang, X.; Yang, J.; Lei, P.; Wen, T. LncRNA MALAT1 shuttled by bone marrow-derived mesenchymal stem cells-secreted exosomes alleviates osteoporosis through mediating microRNA-34c/SATB2 axis. Aging 2019, 11, 8777–8791. [Google Scholar] [CrossRef]
- Zampetaki, A.; Mayr, M. Analytical challenges and technical limitations in assessing circulating MiRNAs. Thromb. Haemost. 2012, 108, 592–598. [Google Scholar] [PubMed] [Green Version]
- Cheng, H.H.; Yi, H.S.; Kim, Y.; Kroh, E.M.; Chien, J.W.; Eaton, K.D.; Goodman, M.T.; Tait, J.F.; Tewari, M.; Pritchard, C.C. Plasma Processing Conditions Substantially Influence Circulating microRNA Biomarker Levels. PLoS ONE 2013, 8, e64795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.J.; Linnstaedt, S.; Palma, J.; Park, J.C.; Ntrivalas, E.; Kwak-Kim, J.Y.; Gilman-Sachs, A.; Beaman, K.; Hastings, M.L.; Martin, J.N.; et al. Plasma components affect accuracy of circulating cancer-related microRNA quantitation. J. Mol. Diagn. 2012, 14, 71–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroh, E.M.; Parkin, R.K.; Mitchell, P.S.; Tewari, M. Analysis of circulating microRNA biomarkers in plasma and serum using quantitative reverse transcription-PCR (qRT-PCR). Methods 2010, 50, 298–301. [Google Scholar] [CrossRef] [Green Version]
- Witwer, K.W. XenomiRs and miRNA homeostasis in health and disease: Evidence that diet and dietary miRNAs directly and indirectly influence circulating miRNA profiles. RNA Biol. 2012, 9, 1147–1154. [Google Scholar] [CrossRef]
- Shende, V.R.; Goldrick, M.M.; Ramani, S.; Earnest, D.J. Expression and Rhythmic Modulation of Circulating MicroRNAs Targeting the Clock Gene Bmal1 in Mice. PLoS ONE 2011, 6, e22586. [Google Scholar] [CrossRef] [Green Version]
- Baggish, A.L.; Hale, A.; Weiner, R.B.; Lewis, G.D.; Systrom, D.; Wang, F.; Wang, T.J.; Chan, S.Y. Dynamic regulation of circulating microRNA during acute exhaustive exercise and sustained aerobic exercise training. J. Physiol. 2011, 589, 3983–3994. [Google Scholar] [CrossRef]
- Sourvinou, I.S.; Markou, A.; Lianidou, E.S. Quantification of circulating miRNAs in plasma: Effect of preanalytical and analytical parameters on their isolation and stability. J. Mol. Diagn. 2013, 15, 827–834. [Google Scholar] [CrossRef]
- Mestdagh, P.; Hartmann, N.; Baeriswyl, L.; Andreasen, D.; Bernard, N.; Chen, C.; Cheo, D.; D’Andrade, P.; DeMayo, M.; Dennis, L.; et al. Evaluation of quantitative mirnA expression platforms in the micrornA quality control (mirQC) study. Nat. Methods 2014, 11, 809–815. [Google Scholar] [CrossRef]
- Ismail, S.M.; El Boghdady, N.A.; Hamoud, H.S.; Shabayek, M.I. Evaluation of circulating miRNA-208a-3p, miRNA-155–5p and miRNA-637 as potential non-invasive biomarkers and the possible mechanistc insights into pre- and postmenopausal osteoporotic females. Arch. Biochem. Biophys. 2020, 684, 108331. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellavia, D.; Raimondi, L.; Costa, V.; De Luca, A.; Carina, V.; Maglio, M.; Fini, M.; Alessandro, R.; Giavaresi, G. Engineered exosomes: A new promise for the management of musculoskeletal diseases. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 1893–1901. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Li, X.; Zhao, Z.; Qian, J.; Wang, Y.; Cui, J.; Weng, W.; Cao, L.; Chen, X.; Hu, Y.; et al. Reversal of Osteoporotic Activity by Endothelial Cell-Secreted Bone Targeting and Biocompatible Exosomes. Nano Lett. 2019, 19, 3040–3048. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yao, J.; Cai, L.; Liu, T.; Wang, X.; Zhang, Y.; Zhou, Z.; Li, T.; Liu, M.; Lai, R.; et al. Bone-Targeted Extracellular Vesicles from Mesenchymal Stem Cells for Osteoporosis Therapy. Int. J. Nanomed. 2020, 15, 7967–7977. [Google Scholar] [CrossRef]
Figure 1.
Cartoon summarizing the actions of microRNAs (miRNAs) and long non-coding RNAs (lncRNAs) in the cell. The processes of biogenesis and sorting in the cellular compartment as well as the interaction with molecular partners are highlighted. (A) Drosha, a nuclear endonuclease, converts the hairpin-loop RNA primary transcript into a shorter stem-loop structured pre-miRNA. After nuclear export by exportin 5, Dicer cleaves the pre-miRNA stem-loop into an RNA fragment with a two-nucleotide 3′ overhang at each end. Thereafter, the mature miRNA is loaded into the RISC complex by AGO proteins (mainly AGO2). Finally, the target mRNA is sorted and processed for cleavage. (B) LncRNA can work in the nucleus or can be exported into the cytoplasm. In the nucleus, lncRNA can act as (1) signaling, synergistically cooperating with a transcription factor to finely regulate in time and space the gene expression; (2) decoy, undermining a transcription factor; (3) guide, recruiting chromatin-modifying enzymes on a target gene; or (4) scaffold, favoring the formation of a complex or a spatial proximity of two or more proteins. When lncRNA is exported into the cytoplasm, it can sequester a miRNA, acting as a sponge (5), avoiding the assembly of RISC complex and favoring the translation of the mRNA into a protein.
Figure 1.
Cartoon summarizing the actions of microRNAs (miRNAs) and long non-coding RNAs (lncRNAs) in the cell. The processes of biogenesis and sorting in the cellular compartment as well as the interaction with molecular partners are highlighted. (A) Drosha, a nuclear endonuclease, converts the hairpin-loop RNA primary transcript into a shorter stem-loop structured pre-miRNA. After nuclear export by exportin 5, Dicer cleaves the pre-miRNA stem-loop into an RNA fragment with a two-nucleotide 3′ overhang at each end. Thereafter, the mature miRNA is loaded into the RISC complex by AGO proteins (mainly AGO2). Finally, the target mRNA is sorted and processed for cleavage. (B) LncRNA can work in the nucleus or can be exported into the cytoplasm. In the nucleus, lncRNA can act as (1) signaling, synergistically cooperating with a transcription factor to finely regulate in time and space the gene expression; (2) decoy, undermining a transcription factor; (3) guide, recruiting chromatin-modifying enzymes on a target gene; or (4) scaffold, favoring the formation of a complex or a spatial proximity of two or more proteins. When lncRNA is exported into the cytoplasm, it can sequester a miRNA, acting as a sponge (5), avoiding the assembly of RISC complex and favoring the translation of the mRNA into a protein.

Figure 2.
Schematization of the biogenesis and formation of the main classes of extracellular vesicles (EVs) released from a cell. (A) Early endosomes involved in the sorting and recycling of canonical intracellular vesicles can generate a multivesicular body (MVB), entrapping vesicles. An MVB can fuse with plasma membrane releasing its content in extracellular space. (B) Plasma membrane can undergo a complex remodeling by sophisticated molecular machinery, generating a bud from which a microvesicle is formed. (C) When a cell experiences a severe injury triggering irreversible damage, the cell activates the apoptotic pathway implying the organized dismantling of the cytoplasm. This process induces the release of the apoptotic bodies. sEVs = small extracellular vesicles; m/lEVs = medium/large extracellular vesicles.
Figure 2.
Schematization of the biogenesis and formation of the main classes of extracellular vesicles (EVs) released from a cell. (A) Early endosomes involved in the sorting and recycling of canonical intracellular vesicles can generate a multivesicular body (MVB), entrapping vesicles. An MVB can fuse with plasma membrane releasing its content in extracellular space. (B) Plasma membrane can undergo a complex remodeling by sophisticated molecular machinery, generating a bud from which a microvesicle is formed. (C) When a cell experiences a severe injury triggering irreversible damage, the cell activates the apoptotic pathway implying the organized dismantling of the cytoplasm. This process induces the release of the apoptotic bodies. sEVs = small extracellular vesicles; m/lEVs = medium/large extracellular vesicles.

Figure 3.
Intercellular exchanges of non-coding RNAs by means of EVs among bone-resident cells. Non-coding RNAs exerting osteopenic activity (increase in osteoclastogenesis or decrease in osteogenesis) are reported. Only ncRNAs directly found encompassed in EVs are reported in the figures. The arrows indicate the origin-to-destination direction between cells (donor to target cell). 1: soluble myostatin, released by aging muscle, affects osteocytes in a paracrine manner, reacting with the drop in endogenous miR-218a (↓miR-218a). The miR-218a-poor EVs released by the affected osteocytes are taken up by osteoblasts, contributing to the perturbation of osteogenesis.
Figure 3.
Intercellular exchanges of non-coding RNAs by means of EVs among bone-resident cells. Non-coding RNAs exerting osteopenic activity (increase in osteoclastogenesis or decrease in osteogenesis) are reported. Only ncRNAs directly found encompassed in EVs are reported in the figures. The arrows indicate the origin-to-destination direction between cells (donor to target cell). 1: soluble myostatin, released by aging muscle, affects osteocytes in a paracrine manner, reacting with the drop in endogenous miR-218a (↓miR-218a). The miR-218a-poor EVs released by the affected osteocytes are taken up by osteoblasts, contributing to the perturbation of osteogenesis.


{kind=link}
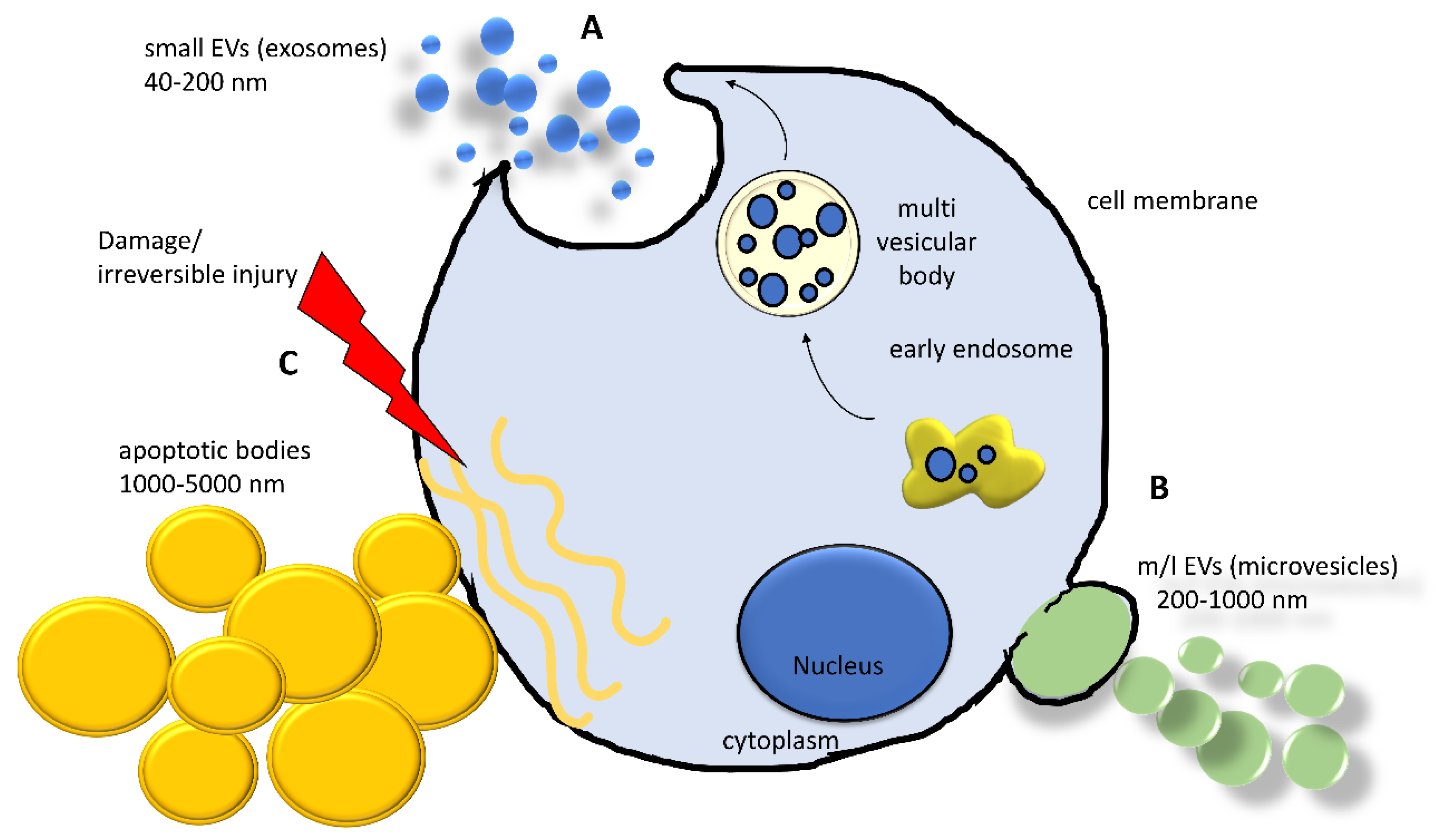{kind=link}
{kind=link}
Table 1.
Intercellular exchange of miRNAs and their osteopenic effects.
| Donor | Mediator | Target | Effect | References |
|---|---|---|---|---|
| Human bone marrow stromal cells | miR-885-5p | Human bone marrow stromal cells | Decrease in osteogenic ability suppressing runx2 and wnt5a | [114] |
| Osteoblasts-like MC3T3 cells | miR-30d-5p, miR-133b, miR-140-3p | Bone marrow stromal cell line ST2 | Inhibition of osteoblast differentiation by runx2 suppression | [115] |
| Dendritic cells, MC3T3 cells, patients’ serum | miR-146a | Osteoblasts, bone marrow mesenchymal stem cells | Induction of cell growth arrest and senescence of osteogenic cells. Increase in RANKL/OPG ratio in osteoblasts. | [117,118,119,120] |
| Adipocytes | miR-138, miR-30c, miR-125a, miR-125b, miR-31 | Mesenchymal stem cells | Suppression of osteocalcin and osteopontin levels | [123] |
| Mononuclear cells, granulocytes | lncRNA JPX | Bone marrow stromal cells | Reduction in alpl, runx2, bglap | [124] |
| Mast cells endothelial cells, | lncRNA MALAT 1 | Macrophages, osteoclasts | Repression of miR-124, inducing the overexpression of mmp9, ctsk, acp5, and car2 | [126,127] |
| Osteoclasts | miR-214 | Osteoblasts | Inhibition of osteoblast function, sustaining of osteoporosis in OVX mouse model | [128] |
| Endothelial cells | miR-31 | Mesenchymal stem cells | Inhibition of the osteogenic differentiation by suppression of Frizzled-3 | [137] |
| Mouse myoblasts | miR-34a | Bone marrow stromal cells | Induction of Sirtuin 1 reduction and senescence in bone marrow stromal cells | [143] |
| Myoblasts | Myostatin/miR-218 | Osteocytic cells Ocy454 | Decrease in osteocytic miR-218 leading to an increase in RANKL expression and decrease in SOST. | [147] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Muraca, M.; Cappariello, A. The Role of Extracellular Vesicles (EVs) in the Epigenetic Regulation of Bone Metabolism and Osteoporosis. Int. J. Mol. Sci. 2020, 21, 8682. https://doi.org/10.3390/ijms21228682
AMA Style
Muraca M, Cappariello A. The Role of Extracellular Vesicles (EVs) in the Epigenetic Regulation of Bone Metabolism and Osteoporosis. International Journal of Molecular Sciences. 2020; 21(22):8682. https://doi.org/10.3390/ijms21228682
Chicago/Turabian StyleMuraca, Maurizio, and Alfredo Cappariello. 2020. "The Role of Extracellular Vesicles (EVs) in the Epigenetic Regulation of Bone Metabolism and Osteoporosis" International Journal of Molecular Sciences 21, no. 22: 8682. https://doi.org/10.3390/ijms21228682
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.