
Anti-Apoptotic Effect of Apelin in Human Placenta: Studies on BeWo Cells and Villous Explants from Third-Trimester Human Pregnancy

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effect of Apelin on mRNA and Protein Expression of Apoptotic Factors in BeWo Cells
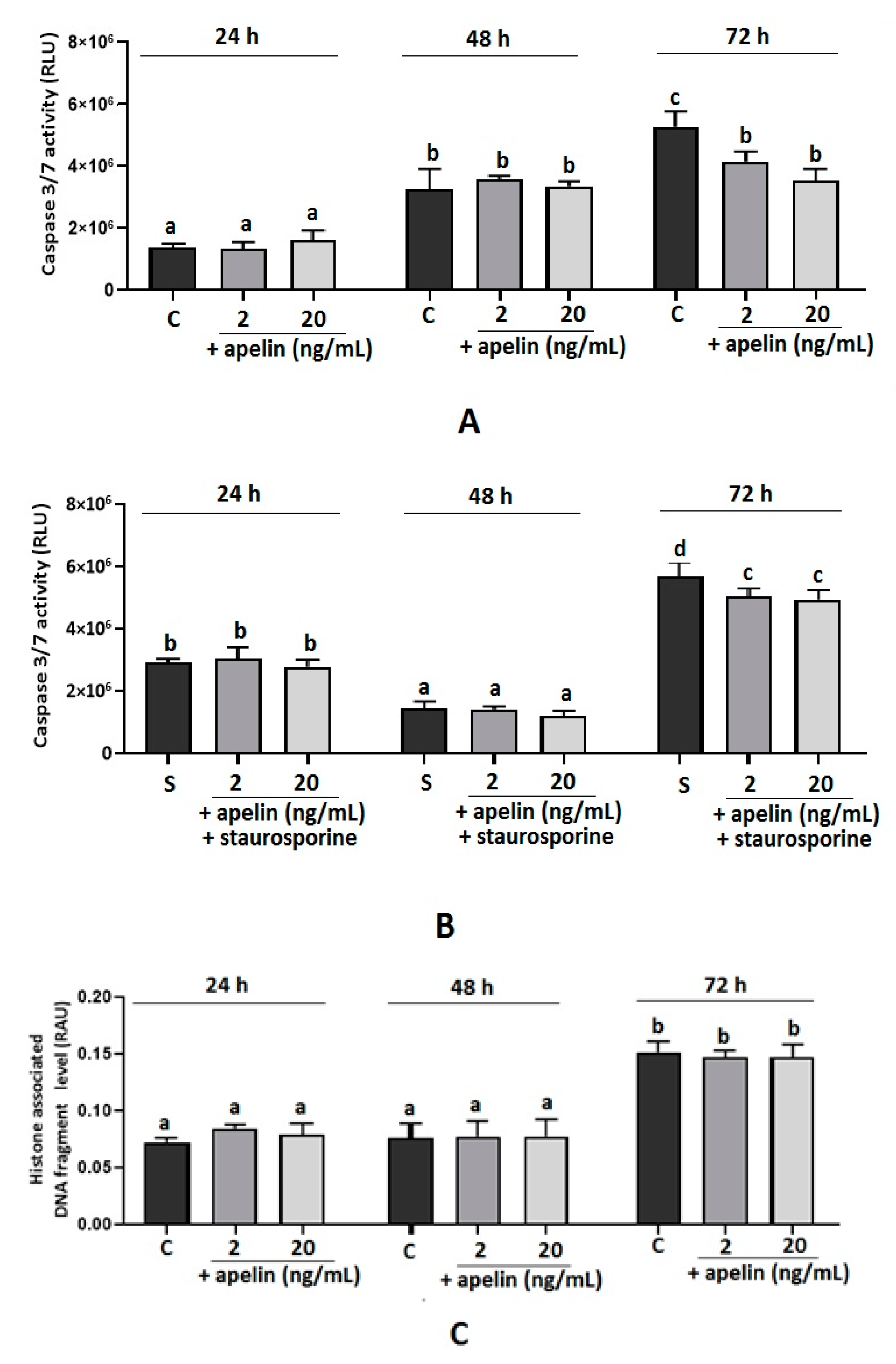
2.2. Effect of Apelin on Caspase 3/7 Activity and DNA Fragmentation in BeWo Cells
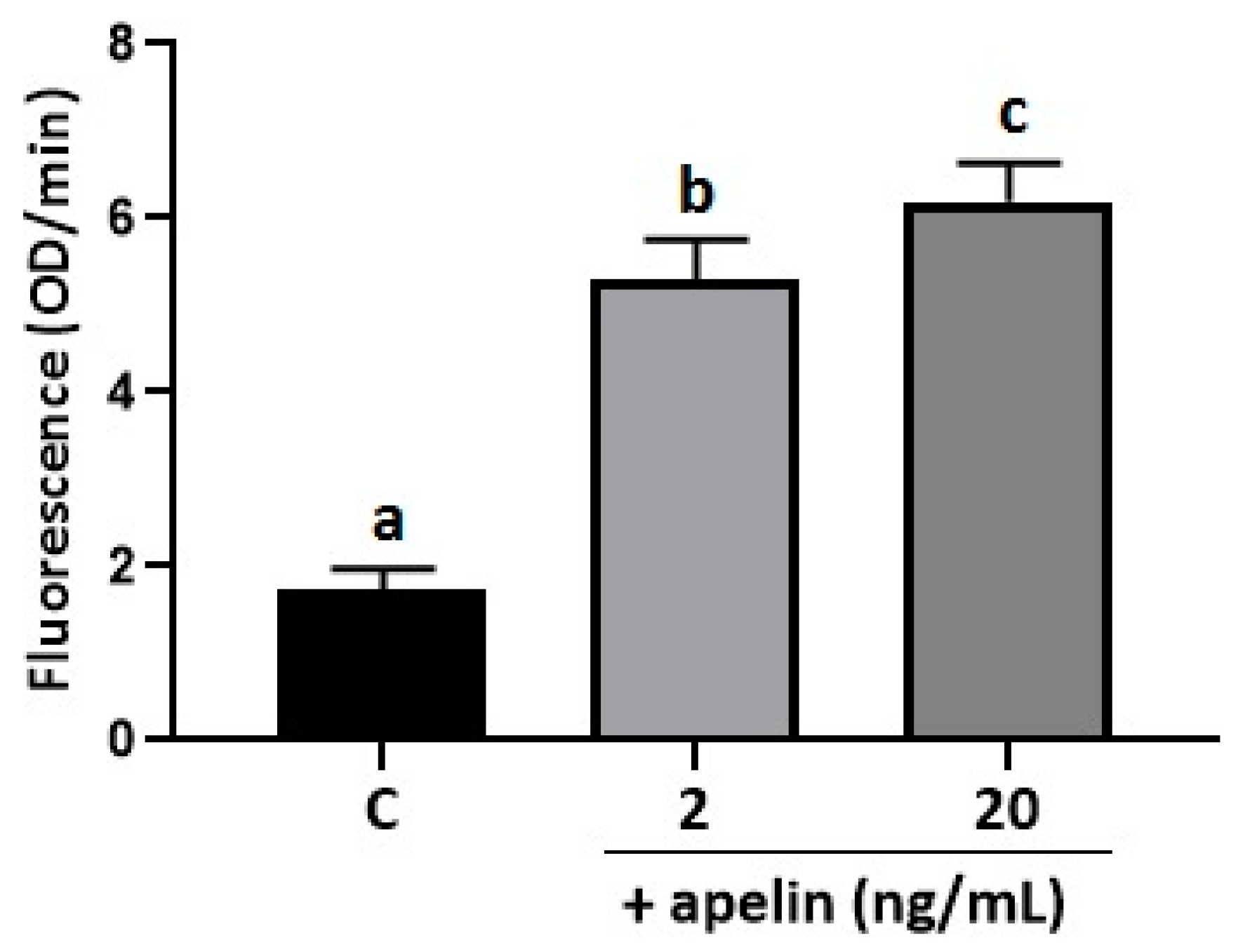
2.3. Effect of Apelin on Oxidative Stress in BeWo Cells
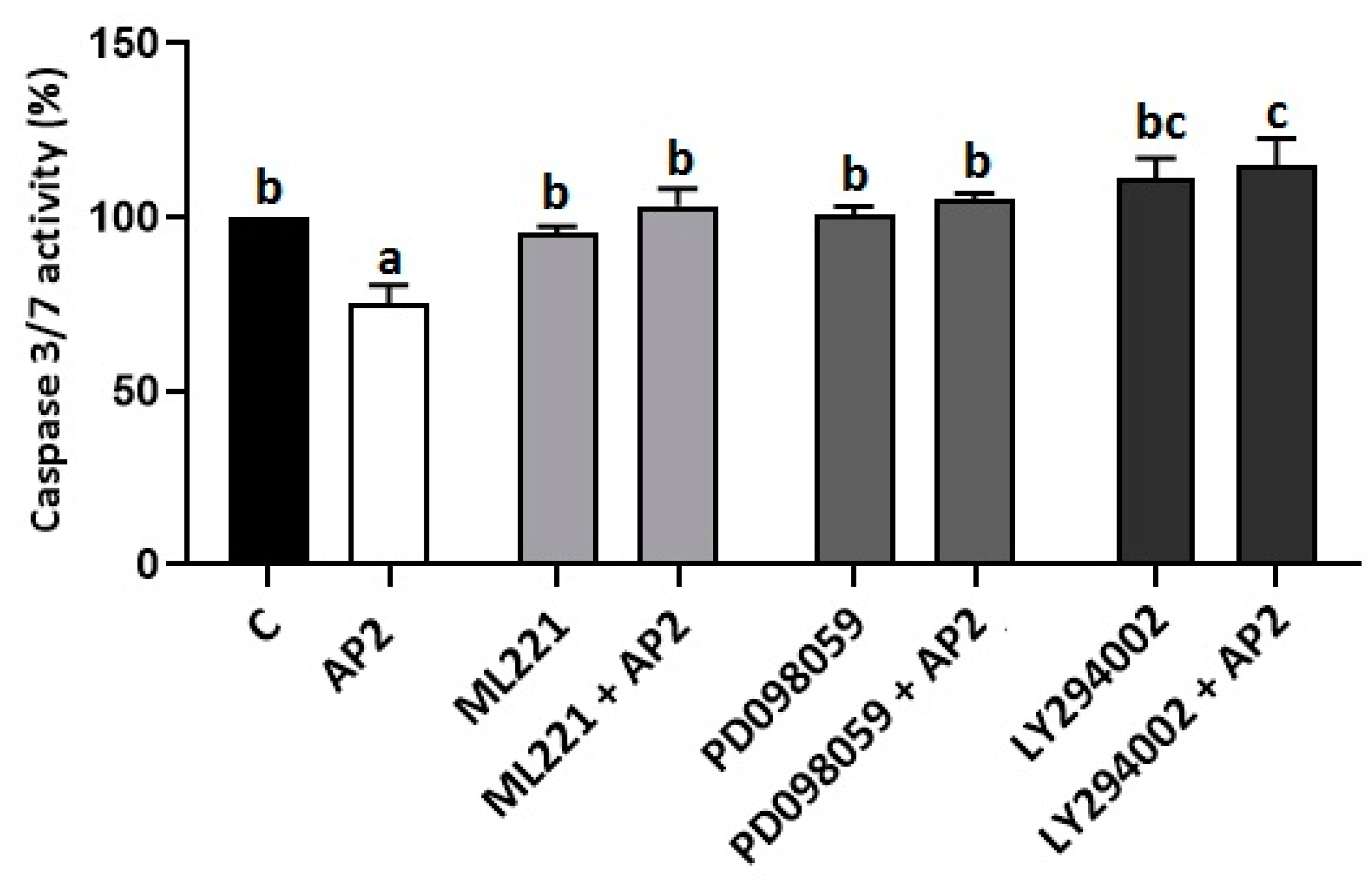
2.4. Involvement of the APJ Receptor and MAP3/1 Kinase in Antiapoptotic Effect of Apelin in BeWo Cells
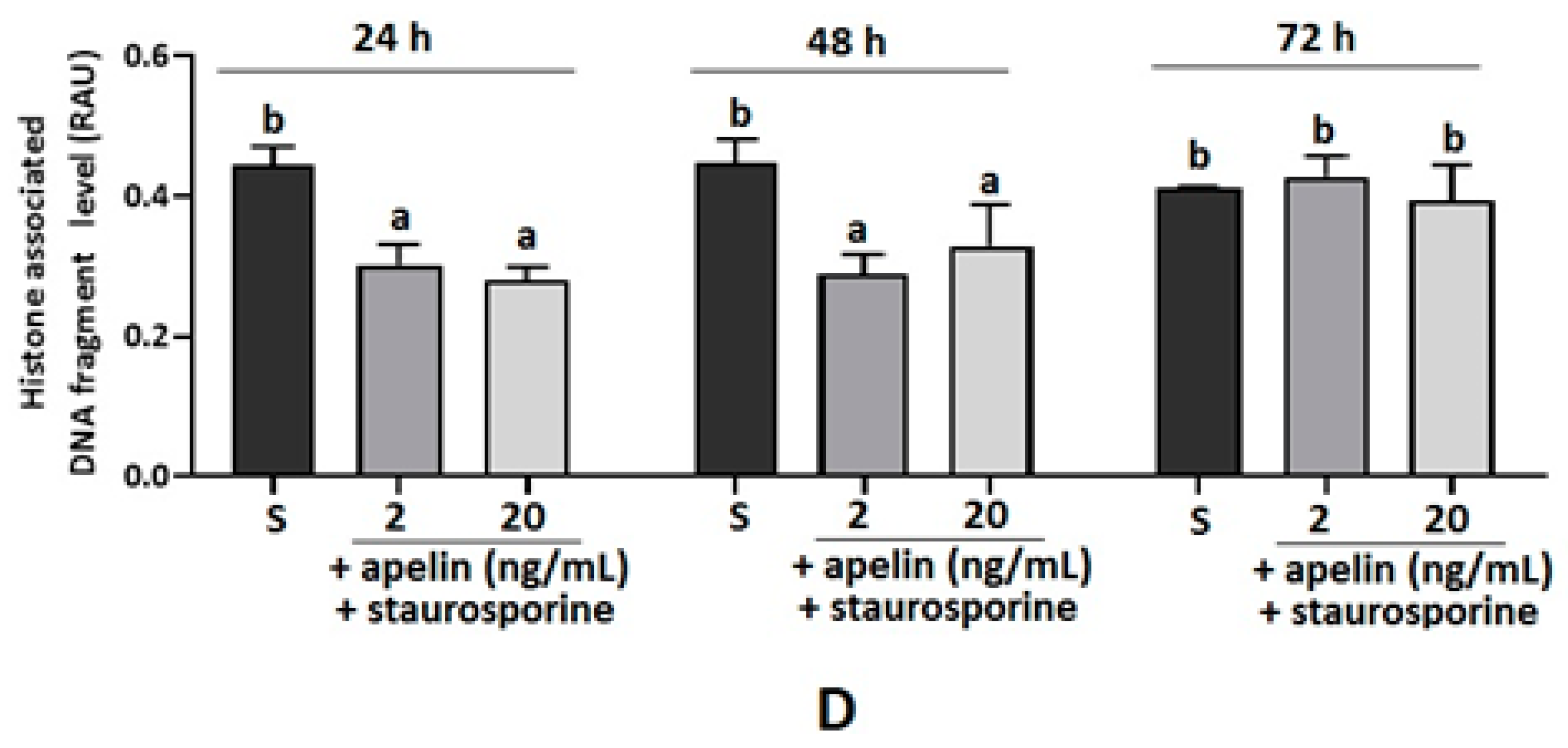
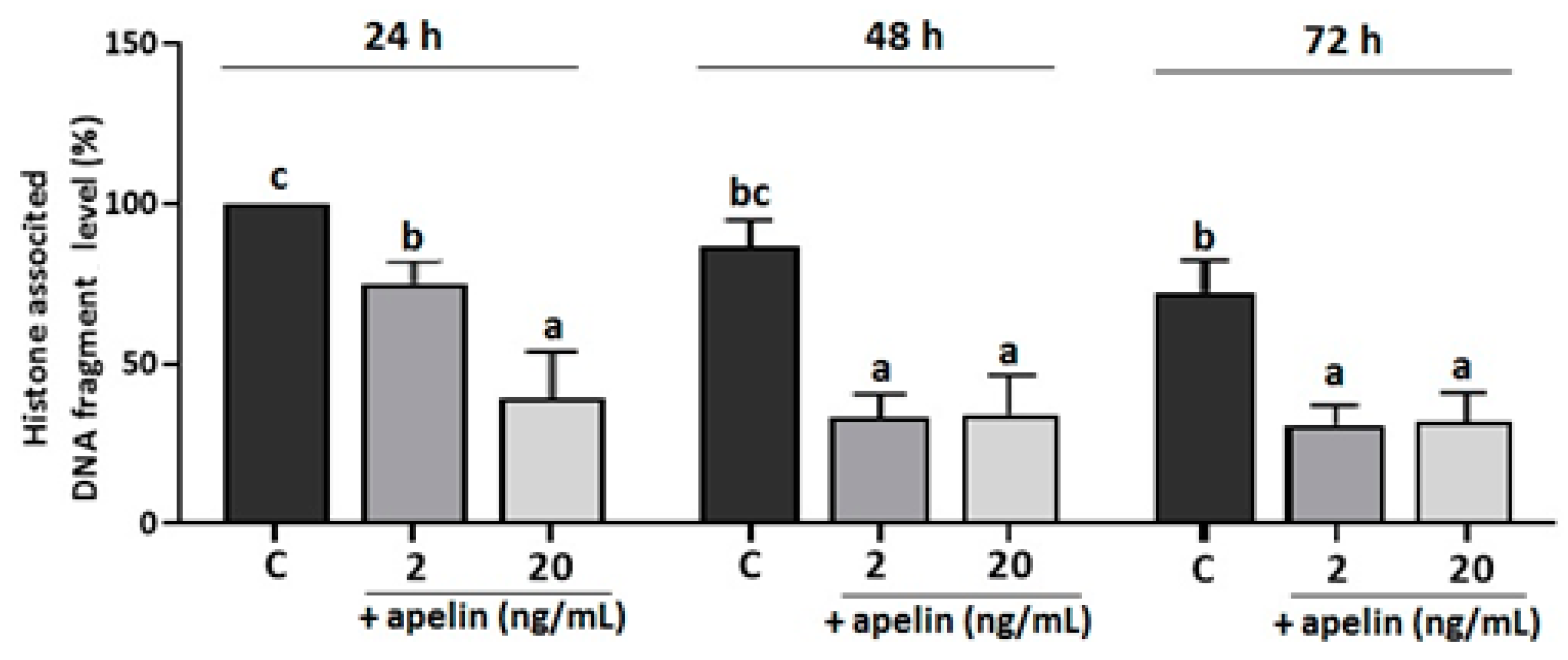
2.5. Effect of Apelin on DNA Fragmentation in Villous Explants from the Third Trimester of Human Pregnancy
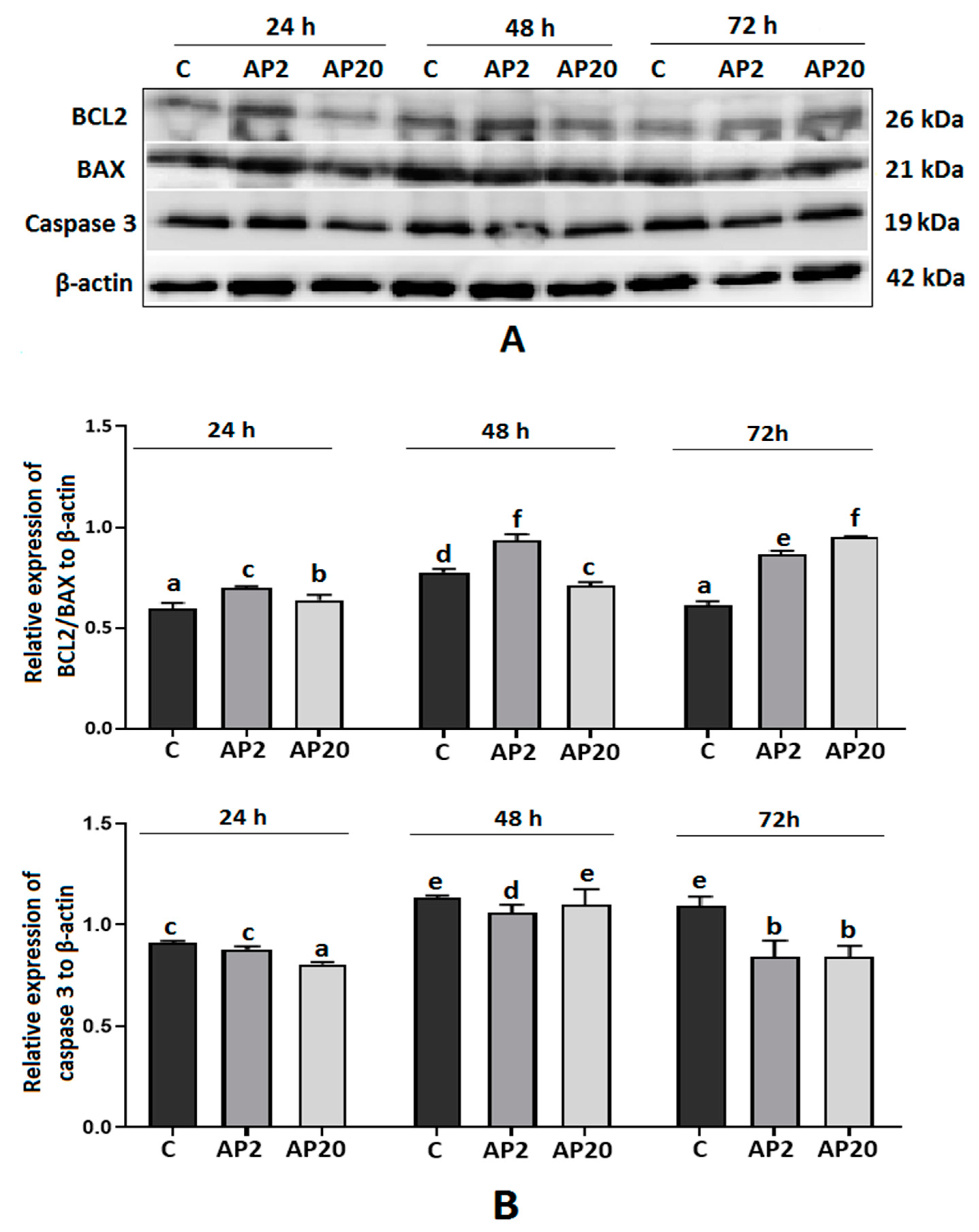
2.6. Effect of Apelin on Protein Expression of BCL2, BAX and Caspase-3 in Villous Explants from the Third Trimester of Human Pregnancy
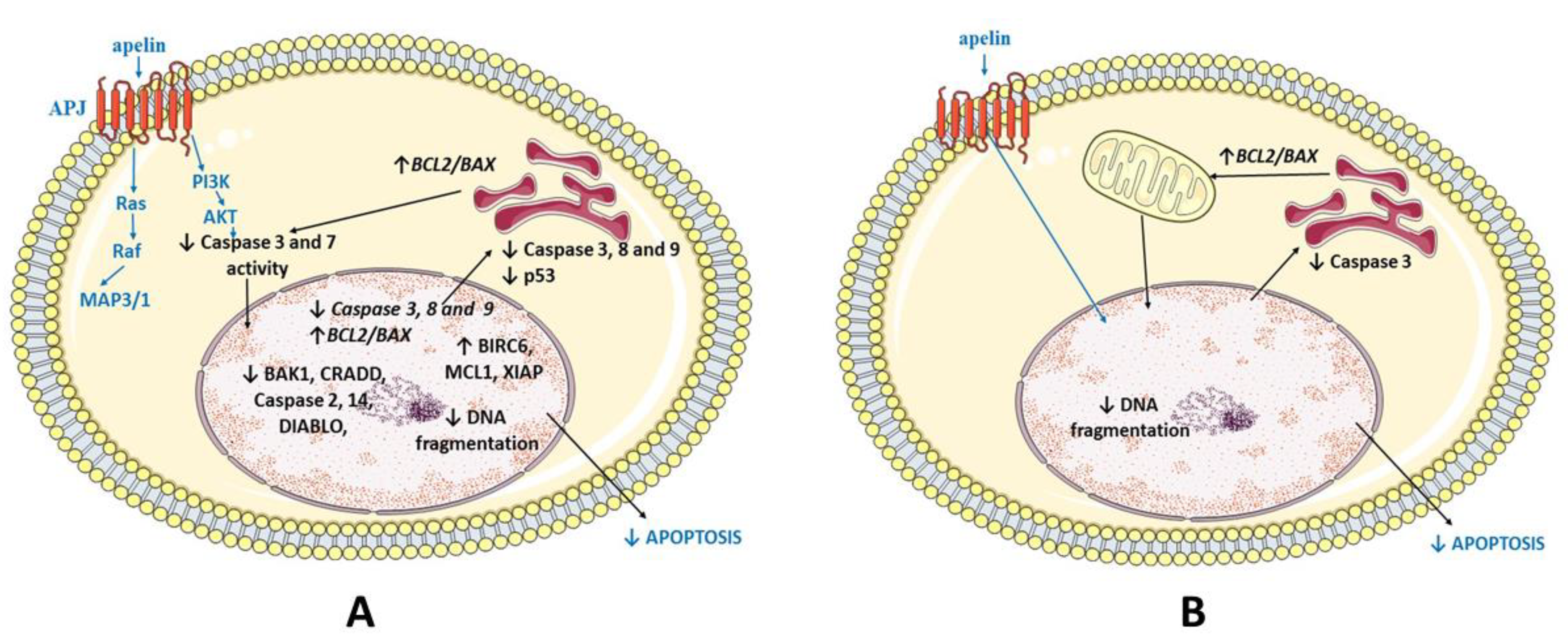
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. In Vitro Culture of BeWo Cells
4.3. In Vitro Culture of Villous Explants from Human Placenta
4.4. Experimental Procedure
4.5. Real-Time PCR
4.6. Western Blot Analysis
4.7. Extracellular Consumption Assay
4.8. Caspase-Glo 3/7 Assay
4.9. Cell Death Detection ELISA Kit
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AKT | Protein kinase B |
| AP2 | Apelin at dose 2 ng/mL |
| AP20 | Apelin at dose 20 ng/mL |
| APAF1 | Apoptotic protease-activating factor 1 |
| BAK | Bcl-2 homologous antagonist/killer |
| BAX | Bcl-2-like protein 4 |
| BCL2 | B-cell lymphoma 2 |
| BCL3 | B-cell lymphoma 3 protein |
| BID | BH3-interacting domain death agonist |
| BIRC6 | Baculoviral IAP repeat-containing 6 |
| BOK | Bcl-2-related ovarian killer protein |
| BSA | Bovine Serum Albumin |
| CASP 7 | Caspase 7 |
| CASP14 | Caspase 14 |
| CASP2 | Caspase 2 |
| CASP3 | Caspase 3 |
| CASP8 | Caspase 8 |
| CRADD | Caspase and RIP adapter with death domain |
| CST | Cell Signaling Technology |
| Ct | Cycle threshold number |
| DAB | 3,3′-diaminobenzidine |
| DEDD | Death effector domain containing |
| DIABLO | Diablo IAP-binding mitochondrial protein |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| E2 | Oestradiol |
| EDTA | ethylenediaminetetraacetic acid |
| ERK1/2/MAP3/1 | Mitogen-activated protein kinase |
| FBS | Fetal Bovine Serum |
| hCG | Chorionic gonadotropin |
| hPL | Human placental lactogen |
| IUGR | Intrauterine growth restriction |
| MCL-1 | Induced myeloid leukemia cell differentiation protein MCL1 |
| NOD1 | Nucleotide-binding oligomerization domain containing protein 1 |
| P4 | Progesterone |
| PBS | Phosphate-buffered saline |
| pyr-apelin-13 | Pyroglutamylated apelin-13 |
| RAU | Relative Absorbance Unit |
| RLU | Relative Luminescence Unit |
| ROS | Reactive oxygen species |
| SDS | sodium dodecyl sulfate |
| SEM | Standard error of the mean |
| TNFR | Tumor necrosis factor receptor |
| TNFRSF25 | Tumor necrosis factor receptor superfamily member 25 |
| XIAP | X-linked inhibitor of apoptosis |
References
- Maltepe, E.; Bakardjiev, A.I.; Fisher, S.J. The Placenta: Transcriptional, Epigenetic, and Physiological Integration during Development. J. Clin. Investig. 2010, 120, 1016–1025. [Google Scholar] [CrossRef] [PubMed]
- Sharp, A.N.; Heazell, A.E.P.; Crocker, I.P.; Mor, G. Placental Apoptosis in Health and Disease: Placental apoptosis in health and disease. Am. J. Reprod. Immunol. 2010, 64, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Cory, S.; Adams, J.M. The Bcl2 Family: Regulators of the Cellular Life-or-Death Switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Arends, M.J.; Wyllie, A.H. Apoptosis: Mechanisms and Roles in Pathology. Int. Rev. Exp. Pathol. 1991, 32, 223–254. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.C.; Leung, T.N.; To, K.F.; Baker, P.N. Apoptosis Is a Rare Event in First-Trimester Placental Tissue. Am. J. Obstet. Gynecol. 2000, 183, 697–699. [Google Scholar] [CrossRef] [PubMed]
- Halperin, R.; Peller, S.; Rotschild, M.; Bukovsky, I.; Schneider, D. Placental Apoptosis in Normal and Abnormal Pregnancies. Gynecol. Obstet. Investig. 2000, 50, 84–87. [Google Scholar] [CrossRef]
- Straszewski-Chavez, S.L.; Abrahams, V.M.; Mor, G. The Role of Apoptosis in the Regulation of Trophoblast Survival and Differentiation during Pregnancy. Endocr. Rev. 2005, 26, 877–897. [Google Scholar] [CrossRef] [Green Version]
- Soni, S.; Rath, G.; Prasad, C.P.; Salhan, S.; Saxena, S.; Jain, A.K. Apoptosis and Bcl-2 Protein Expression in Human Placenta over the Course of Normal Pregnancy: Apoptosis and Bcl-2 in Normal Human Placenta. Anat. Histol. Embryol. 2010, 39, 426–431. [Google Scholar] [CrossRef]
- Ishihara, N.; Matsuo, H.; Murakoshi, H.; Laoag-Fernandez, J.B.; Samoto, T.; Maruo, T. Increased Apoptosis in the Syncytiotrophoblast in Human Term Placentas Complicated by Either Preeclampsia or Intrauterine Growth Retardation. Am. J. Obstet. Gynecol. 2002, 186, 158–166. [Google Scholar] [CrossRef]
- Erel, C.T.; Dane, B.; Calay, Z.; Kaleli, S.; Aydinli, K. Apoptosis in the Placenta of Pregnancies Complicated with IUGR. Int. J. Gynaecol. Obstet. 2001, 73, 229–235. [Google Scholar] [CrossRef]
- Dagdelen, M.; Temur, M.; Yılmaz, Ö.; Altındag, T.; Uslu, T.; Özbay, P.Ö. Placental Bed Apoptosis Is Increased in Pregnant Women with Pre-Eclampsia versus Normotensive Pregnant Women. J. Obstet. Gynaecol. 2016, 36, 974–979. [Google Scholar] [CrossRef]
- Kelten, C.; Zekioglu, O.; Terek, C.; Ozdemir, N.; Duzcan, E. Expression of Ki-67, Bcl-2 and Bax in the First Trimester Abortion Materials. Turk. Patoloji. Derg. 2010, 26, 31. [Google Scholar] [CrossRef] [Green Version]
- Ishihara, N.; Matsuo, H.; Murakoshi, H.; Laoag-Fernandez, J.; Samoto, T.; Maruo, T. Changes in Proliferative Potential, Apoptosis and Bcl-2 Protein Expression in Cytotrophoblasts and Syncytiotrophoblast in Human Placenta over the Course of Pregnancy. Endocr. J. 2000, 47, 317–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.C.; Baker, P.N.; Symonds, E.M. Increased placental apoptosis in intrauterine growth restriction. Am. J. Obstet. Gynecol. 1997, 177, 1395–1401. [Google Scholar] [CrossRef]
- Pérez-Pérez, A.; Toro, A.R.; Vilarino-Garcia, T.; Guadix, P.; Maymó, J.L.; Dueñas, J.L.; Varone, C.L.; Sánchez-Margalet, V. Leptin Reduces Apoptosis Triggered by High Temperature in Human Placental Villous Explants: The Role of the P53 Pathway. Placenta 2016, 42, 106–113. [Google Scholar] [CrossRef]
- Toro, A.R.; Maymó, J.L.; Ibarbalz, F.M.; Pérez, A.P.; Maskin, B.; Faletti, A.G.; Margalet, V.S.; Varone, C.L. Leptin Is an Anti-Apoptotic Effector in Placental Cells Involving P53 Downregulation. PLoS ONE 2014, 9, e99187. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Pérez, A.; Toro, A.; Vilariño-Garcia, T.; Guadix, P.; Maymó, J.; Dueñas, J.L.; Varone, C.; Sánchez-Margalet, V. Leptin Protects Placental Cells from Apoptosis Induced by Acidic Stress. Cell Tissue Res. 2019, 375, 733–742. [Google Scholar] [CrossRef]
- Duval, F.; Santos, E.D.; Poidatz, D.; Sérazin, V.; Gronier, H.; Vialard, F.; Dieudonné, M.N. Adiponectin Inhibits Nutrient Transporters and Promotes Apoptosis in Human Villous Cytotrophoblasts: Involvement in the Control of Fetal Growth. Biol. Reprod. 2016, 94, 111. [Google Scholar] [CrossRef]
- Jarvenpaa, J.; Vuoristo, J.T.; Santaniemi, M.; Ukkola, O.; Savolainen, E.R.; Jääskeläinen, M.; Tapanainen, J.S.; Kesäniemi, A.; Ryynanen, M. Adiponectin Induced Placental Cell Apoptosis Could Be Mediated via the ADIPOR1-Receptor in Pre-Eclampsia with IUGR. J. Perinat. Med. 2009, 37, 257–262. [Google Scholar] [CrossRef]
- Lee, D.K.; Cheng, R.; Nguyen, T.; Fan, T.; Kariyawasam, A.P.; Liu, Y.; Osmond, D.H.; George, S.R.; O’Dowd, B.F. Characterization of Apelin, the Ligand for the APJ Receptor. J. Neurochem. 2000, 74, 34–41. [Google Scholar] [CrossRef]
- Bertrand, C.; Valet, P.; Castan-Laurell, I. Apelin and Energy Metabolism. Front. Physiol. 2015, 6, 115. [Google Scholar] [CrossRef] [PubMed]
- Kidoya, H.; Takakura, N. Biology of the Apelin-APJ Axis in Vascular Formation. J. Biochem. 2012, 152, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Furuya, M.; Okuda, M.; Usui, H.; Takenouchi, T.; Kami, D.; Nozawa, A.; Shozu, M.; Umezawa, A.; Takahashi, T.; Aoki, I. Expression of Angiotensin II Receptor-like 1 in the Placentas of Pregnancy-Induced Hypertension. Int. J. Gynecol. Pathol. 2012, 31, 227–235. [Google Scholar] [CrossRef]
- Yamaleyeva, L.M.; Chappell, M.C.; Brosnihan, K.B.; Anton, L.; Caudell, D.L.; Shi, S.; McGee, C.; Pirro, N.; Gallagher, P.E.; Taylor, R.N.; et al. Downregulation of Apelin in the Human Placental Chorionic Villi from Preeclamptic Pregnancies. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E852–E860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mlyczyńska, E.; Kurowska, P.; Drwal, E.; Opydo-Chanek, M.; Tworzydło, W.; Kotula-Balak, M.; Rak, A. Apelin and Apelin Receptor in Human Placenta: Expression, Signalling Pathway and Regulation of Trophoblast JEG-3 and BeWo Cells Proliferation and Cell Cycle. Int. J. Mol. Med. 2020, 45, 691–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughan, O.R.; Powell, T.L.; Jansson, T. Apelin Is a Novel Regulator of Human Trophoblast Amino Acid Transport. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E810–E816. [Google Scholar] [CrossRef]
- Mayeur, S.; Wattez, J.S.; Lukaszewski, M.A.; Lecoutre, S.; Butruille, L.; Drougard, A.; Eberlé, D.; Bastide, B.; Laborie, C.; Storme, L.; et al. Apelin Controls Fetal and Neonatal Glucose Homeostasis and Is Altered by Maternal Undernutrition. Diabetes 2016, 65, 554–560. [Google Scholar] [CrossRef] [Green Version]
- Kidoya, H.; Ueno, M.; Yamada, Y.; Mochizuki, N.; Nakata, M.; Yano, T.; Fujii, R.; Takakura, N. Spatial and Temporal Role of the Apelin/APJ System in the Caliber Size Regulation of Blood Vessels during Angiogenesis. EMBO J. 2008, 27, 522–534. [Google Scholar] [CrossRef] [Green Version]
- Dawid, M.; Mlyczyńska, E.; Kurowska, P.; Sierpowski, M.; Rak, A. Apelin Decreased Placental Hormone Secretion by Human Trophoblast BeWo Cells via Apelin Receptor, Protein Kinase A and Extracellular Signal-Regulated Kinases 1/2 Activation. J. Physiol. Pharmacol. 2019. [Google Scholar] [CrossRef]
- Zou, Y.; Wang, B.; Fu, W.; Zhou, S.; Nie, Y.; Tian, S. Apelin-13 Protects PC12 Cells from Corticosterone-Induced Apoptosis Through PI3K and ERKs Activation. Neurochem. Res. 2016, 41, 1635–1644. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, X.; Cui, H.; Zhang, C.; Zhu, C.; Li, L. Apelin-13 Protects the Brain against Ischemia/Reperfusion Injury through Activating PI3K/Akt and ERK1/2 Signaling Pathways. Neurosci. Lett. 2014, 568, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Cui, R.R.; Mao, D.A.; Yi, L.; Wang, C.; Zhang, X.X.; Xie, H.; Wu, X.P.; Liao, X.B.; Zhou, H.; Meng, J.C.; et al. Apelin Suppresses Apoptosis of Human Vascular Smooth Muscle Cells via APJ/PI3-K/Akt Signaling Pathways. Amino Acids 2010, 39, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Foroughi, K.; Khaksari, M.; Rahmati, M.; Bitaraf, F.S.; Shayannia, A. Apelin-13 Protects PC12 Cells Against Methamphetamine-Induced Oxidative Stress, Autophagy and Apoptosis. Neurochem. Res. 2019, 44, 2103–2112. [Google Scholar] [CrossRef]
- Niknazar, S.; Abbaszadeh, H.A.; Peyvandi, H.; Rezaei, O.; Forooghirad, H.; Khoshsirat, S.; Peyvandi, A.A. Protective Effect of [Pyr1]-Apelin-13 on Oxidative Stress-Induced Apoptosis in Hair Cell-like Cells Derived from Bone Marrow Mesenchymal Stem Cells. Eur. J. Pharmacol. 2019, 853, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lin, X.; Xu, J.; Tang, F. Apelin-13 Reduces Oxidative Stress Induced by Uric Acid via Downregulation of Renin-Angiotensin System in Adipose Tissue. Toxicol. Lett. 2019, 305, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Van Mieghem, T.; van Bree, R.; Van Herck, E.; Pijnenborg, R.; Deprest, J.; Verhaeghe, J. Maternal Apelin Physiology during Rat Pregnancy: The Role of the Placenta. Placenta 2010, 31, 725–730. [Google Scholar] [CrossRef]
- Cobellis, L.; De Falco, M.; Mastrogiacomo, A.; Giraldi, D.; Dattilo, D.; Scaffa, C.; Colacurci, N.; De Luca, A. Modulation of Apelin and APJ Receptor in Normal and Preeclampsia-Complicated Placentas. Histol. Histopathol. 2007, 22, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Alipour, F.G.; Ashoori, M.R.; Pilehvar-Soltanahmadi, Y.; Zarghami, N. An Overview on Biological Functions and Emerging Therapeutic Roles of Apelin in Diabetes Mellitus. Diabetes Metab. Syndr. 2017, 11, S919–S923. [Google Scholar] [CrossRef] [PubMed]
- Eberlé, D.; Marousez, L.; Hanssens, S.; Knauf, C.; Breton, C.; Deruelle, P.; Lesage, J. Elabela and Apelin Actions in Healthy and Pathological Pregnancies. Cytokine Growth Factor Rev. 2019, 46, 45–53. [Google Scholar] [CrossRef]
- Atia, T.A. Placental Apoptosis in Recurrent Miscarriage. Kaohsiung J. Med. Sci. 2017, 33, 449–452. [Google Scholar] [CrossRef] [Green Version]
- Harris, L.K.; Pantham, P.; Yong, H.E.J.; Pratt, A.; Borg, A.J.; Crocker, I.; Westwood, M.; Aplin, J.; Kalionis, B.; Murthi, P. The Role of Insulin-like Growth Factor 2 Receptor-Mediated Homeobox Gene Expression in Human Placental Apoptosis, and Its Implications in Idiopathic Fetal Growth Restriction. Mol. Hum. Reprod. 2019, 25, 572–585. [Google Scholar] [CrossRef]
- Simenc, J.; Lipnik-Stangelj, M. Staurosporine Induces Different Cell Death Forms in Cultured Rat Astrocytes. Radiol. Oncol. 2012, 46. [Google Scholar] [CrossRef]
- Nardo, T.; Micalizzi, G.; Vicinanza, R.; De Iuliis, F.; Taglieri, L.; Scarpa, S. Adhesion to Type V Collagen Enhances Staurosporine-Induced Apoptosis of Adrenocortical Cancer Cells. Tumor Biol. 2014, 35, 9949–9955. [Google Scholar] [CrossRef]
- Sokolova, Y.Y.; Bowers, L.C.; Alvarez, X.; Didier, E.S. Encephalitozoon Cuniculi and Vittaforma Corneae (Phylum Microsporidia) Inhibit Staurosporine-Induced Apoptosis in Human THP-1 Macrophages in Vitro. Parasitology 2019, 146, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Aminyavari, S.; Zahmatkesh, M.; Farahmandfar, M.; Khodagholi, F.; Dargahi, L.; Zarrindast, M.R. Protective Role of Apelin-13 on Amyloid Β25–35-Induced Memory Deficit; Involvement of Autophagy and Apoptosis Process. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 89, 322–334. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Gao, W.; Shan, X.; Wang, C.; Wang, H.; Shao, Z.; Dou, S.; Jiang, Y.; Wang, C.; Cheng, B. Apelin-36 Mediates Neuroprotective Effects by Regulating Oxidative Stress, Autophagy and Apoptosis in MPTP-Induced Parkinson’s Disease Model Mice. Brain Res. 2020, 1726, 146493. [Google Scholar] [CrossRef] [PubMed]
- Shuang, L.; Jidong, W.; Hongjuan, P.; Zhenwei, Y. Effects of Apelin on Proliferation and Apoptosis in Rat Ovarian Granulosa Cells. Clin. Exp. Obstet. Gynecol. 2016, 43, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Tian, F.J.; Lin, Y. Oxidative Stress in Placenta: Health and Diseases. BioMed Res. Int. 2015, 2015, 293271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kingdom, J.C.P.; Kaufmann, P. Oxygen and Placental Villous Development: Origins of Fetal Hypoxia. Placenta 1997, 18, 613–621. [Google Scholar] [CrossRef]
- Khera, A.; Vanderlelie, J.J.; Perkins, A.V. Selenium supplementation protects trophoblast cells from mitochondrial oxidative stress. Placenta 2013, 34, 594–598. [Google Scholar] [CrossRef] [Green Version]
- Cross, T.G.; Scheel-Toellner, D.; Henriquez, N.V.; Deacon, E.; Salmon, M.; Lord, J.M. Serine/Threonine Protein Kinases and Apoptosis. Exp. Cell Res. 2000, 256, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Devine, D.; Sterner, J.; Parker, P.; Basen-Engquist, K.; de Moor, C.; Fitzgerald, M.; Cohen, L. Social aspects of religiosity predict reductions in anxiety for women undergoing treatment for ovarian cancer. Psychosom. Med. 2002, 64, 122. [Google Scholar]
- Fruehauf, J.P. In Vitro Assay-Assisted Treatment Selection for Women with Breast or Ovarian Cancer. Endocr. Relat. Cancer 2002, 9, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Blandau, R.; Warrick, E.; Rumery, R.E. In Vitro Cultivation of Fetal Mouse Ovaries. Fertil. Steril. 1965, 16, 705–715. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Konieczka, P.; Barszcz, M.; Kowalczyk, P.; Szlis, M.; Jankowski, J. The potential of acetylsalicylic acid and vitamin E in modulating inflammatory cascades in chickens under lipopolysaccharide-induced inflammation. Vet. Res. 2019, 50, 1–10. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay ID | Description of Gene | Gene Symbol | Average Fold Change | |
|---|---|---|---|---|
| AP2 | AP20 | |||
| Pro-Survival Protein | ||||
| Hs00608023_m1 | B-cell lymphoma 2 | BCL2 | 0.26 ** | 1.09 |
| Hs00180403_m1 | B-cell lymphoma 3 protein | BCL3 | 0.44 *** | 2.00 ** |
| Hs00212288_m1 | Baculoviral IAP repeat-containing protein 6 | BIRC6 | 1.77 ** | 0.99 |
| Hs00172036_m1 | Induced myeloid leukemia cell differentiation protein Mcl-1 | MCL1 | 1.48 * | 1.12 |
| Hs00745222_s1 | X-linked inhibitor of apoptosis | XIAP | 0.94 | 2.08 *** |
| Pro-Apoptotic Protein | ||||
| Hs00559441_m1 | Apoptotic protease-activating factor 1 (Apaf-1) | APAF1 | 0.93 | 0.77 * |
| Hs00832876_g1 | Bcl-2 homologous antagonist/killer | BAK1 | 0.26 ** | 0.47 ** |
| Hs00751844_s1 | Bcl-2-like protein 4 | BAX | 0.80 * | 0.45 ** |
| Hs00609632_m1 | BH3-interacting domain death agonist | BID | 1.02 | 0.98 |
| Hs00261296_m1 | Bcl-2-related ovarian killer protein | BOK | 0.38 ** | 1.17 |
| Hs00196075_m1 | Nucleotide-binding oligomerization domain containing protein 1 | NOD1 | 0.61 * | 0.60 * |
| Hs01011159_g1 | Caspase and RIP adapter with death domain | CRADD | 0.74 * | 0.94 |
| Hs00201637_m1 | Caspase 14 precursor | CASP14 | 0.58 *** | 0.68 ** |
| Hs00892481_m1 | Caspase 2 precursor | CASP2 | 0.80 *** | 0.51 *** |
| Hs00234387_m1 | Caspase 3 precursor | CASP3 | 0.59 ** | 0.58 ** |
| Hs01018151_m1 | Caspase 8 precursor | CASP8 | 0.59 * | 0.63 * |
| Hs00154260_m1 | Caspase 9 precursor | CASP9 | 1.09 | 0.57 * |
| Hs00172768_m1 | Death effector domain containing | DEDD | 1.12 | 0.97 |
| Hs00219876_m1 | Diablo IAP-binding mitochondrial protein | DIABLO | 0.89 | 0.87 * |
| Hs00980365_g1 | Tumor necrosis factor receptor superfamily member 25 | TNFRSF25 | 0.51 ** | 0.57 * |
| Gene Symbol | Gene Name | Catalog Number | Reference Sequence |
|---|---|---|---|
| BCL2 | B-cell lymphoma 2 | Hs00608023_m1 | NM_000633.2 |
| BCL3 | B-cell lymphoma 3 protein | Hs00180403_m1 | NM_005178.4 |
| BIRC6 | Baculoviral IAP repeat-containing protein 6 | Hs00212288_m1 | NM_016252.3 |
| MCL1 | Induced myeloid leukemia cell differentiation protein Mcl-1 | Hs00172036_m1 | NM_001197320.1 |
| XIAP | X-linked inhibitor of apoptosis | Hs00745222_s1 | NM_001167.3 |
| APAF1 | Apoptotic peptidase-activating factor 1 | Hs00559441_m1 | NM_013229.2 |
| BAK1 | Bcl-2 homologous antagonist/killer | Hs00832876_g1 | NM_001188.3 |
| BAX | Bcl-2-like protein 4 | Hs00751844_s1 | NM_001291428.1 |
| BID | BH3-interacting domain death agonist | Hs00609632_m1 | NM_197966.2 |
| BOK | Bcl-2-related ovarian killer protein | Hs00261296_m1 | NM_032515.4 |
| NOD1 | Nucleotide-binding oligomerization domain containing protein 1 | Hs00196075_m1 | NM_006092.2 |
| CRADD | Caspase and RIP adapter with death domain | Hs01011159_g1 | NM_001320099.1 |
| CASP14 | Caspase-14 precursor | Hs00201637_m1 | NM_012114.2 |
| CASP2 | Caspase-2 precursor | Hs00892481_m1 | NM_001224.4 |
| CASP3 | Caspase-3 precursor | Hs00234387_m1 | NM_004346.3 |
| CASP8 | Caspase-8 precursor | Hs01018151_m1 | NM_001228.4 |
| CASP9 | Caspase-9 precursor | Hs00154260_m1 | NM_001229.4 |
| DEDD | Death effector domain containing | Hs00172768_m1 | NM_001039711.1 |
| DIABLO | Diablo IAP-Binding Mitochondrial Protein | Hs00219876_m1 | NM_001278302.1 |
| TNFRSF25 | Tumor necrosis factor receptor superfamily member 25 | Hs00980365_g1 | NM_003790.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mlyczyńska, E.; Myszka, M.; Kurowska, P.; Dawid, M.; Milewicz, T.; Bałajewicz-Nowak, M.; Kowalczyk, P.; Rak, A. Anti-Apoptotic Effect of Apelin in Human Placenta: Studies on BeWo Cells and Villous Explants from Third-Trimester Human Pregnancy. Int. J. Mol. Sci. 2021, 22, 2760. https://doi.org/10.3390/ijms22052760
Mlyczyńska E, Myszka M, Kurowska P, Dawid M, Milewicz T, Bałajewicz-Nowak M, Kowalczyk P, Rak A. Anti-Apoptotic Effect of Apelin in Human Placenta: Studies on BeWo Cells and Villous Explants from Third-Trimester Human Pregnancy. International Journal of Molecular Sciences. 2021; 22(5):2760. https://doi.org/10.3390/ijms22052760
Chicago/Turabian StyleMlyczyńska, Ewa, Małgorzata Myszka, Patrycja Kurowska, Monika Dawid, Tomasz Milewicz, Marta Bałajewicz-Nowak, Paweł Kowalczyk, and Agnieszka Rak. 2021. "Anti-Apoptotic Effect of Apelin in Human Placenta: Studies on BeWo Cells and Villous Explants from Third-Trimester Human Pregnancy" International Journal of Molecular Sciences 22, no. 5: 2760. https://doi.org/10.3390/ijms22052760