
Effects of the Escherichia coli Bacterial Toxin Cytotoxic Necrotizing Factor 1 on Different Human and Animal Cells: A Systematic Review


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
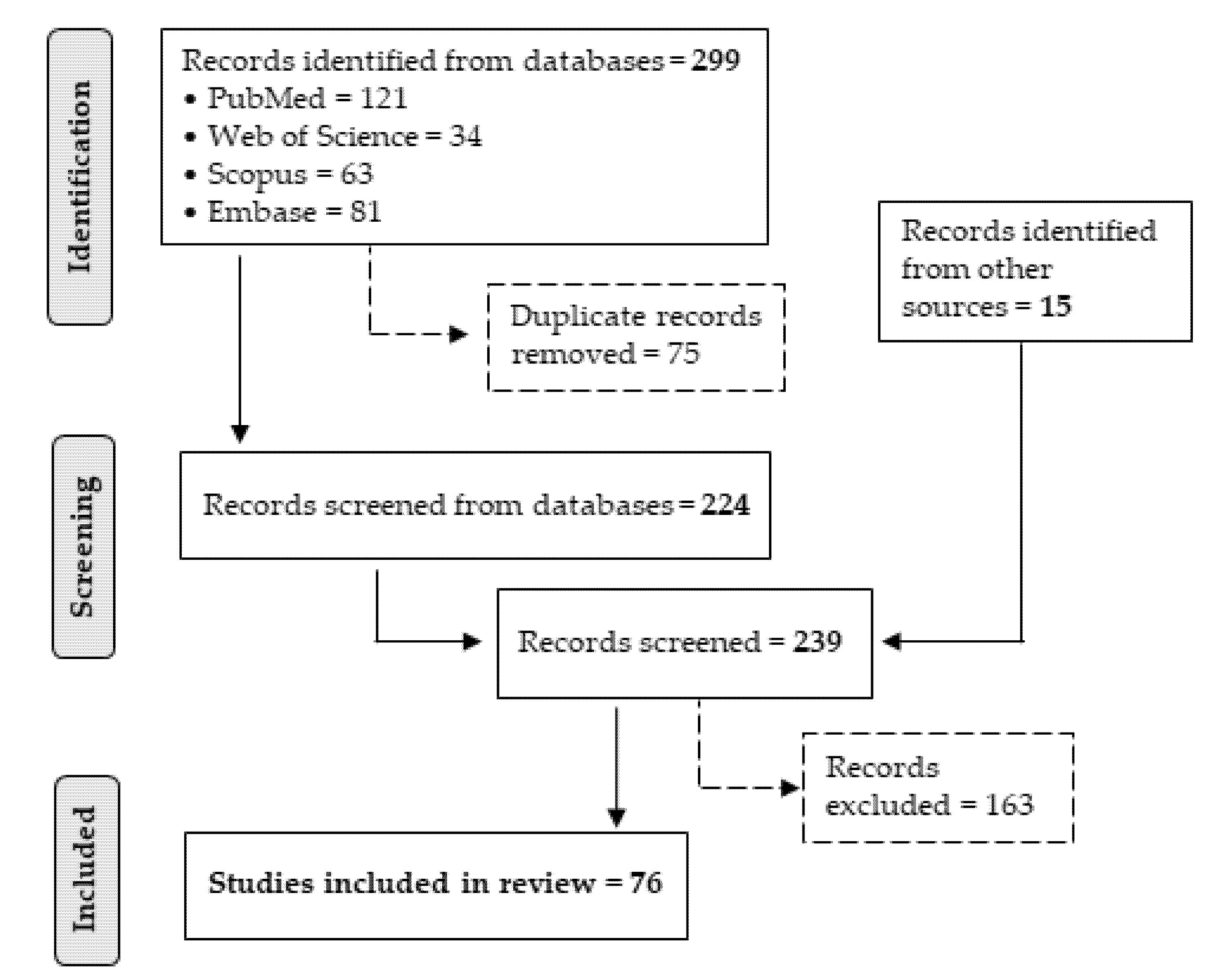
2.1. Literature Search Strategy
2.2. Inclusion Criteria
2.3. Exclusion Criteria
2.4. Data Extraction
3. Results
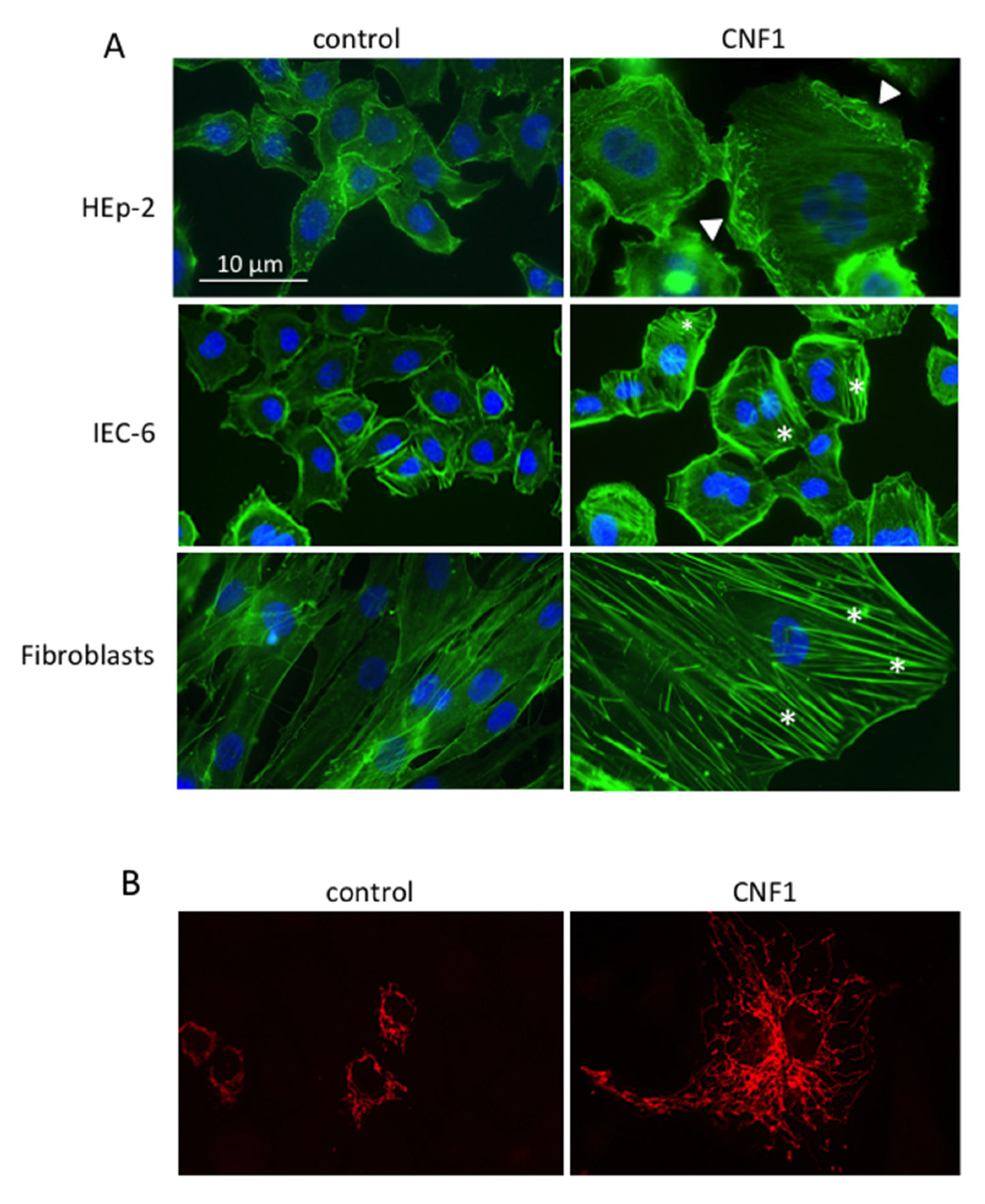
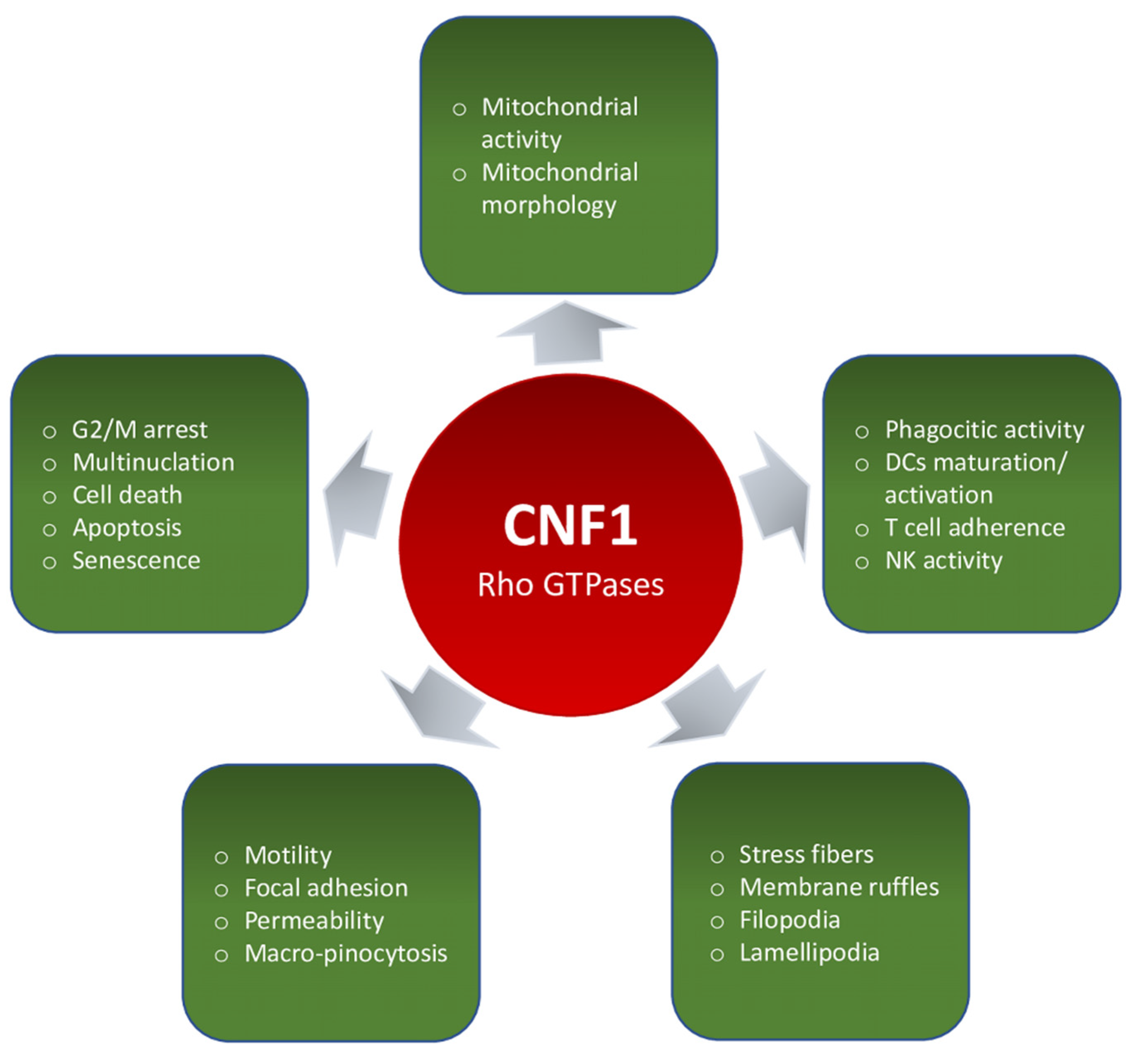
3.1. Effects on Rho GTPases and on Actin Cytoskeleton
3.1.1. Actin Cytoskeleton-Dependent Phenomena (Motility, Focal Adhesion, Permeability, Phago-Pinocytosis)
3.1.2. Multinucleation, Cell Cycle, Cell Death, Apoptosis, and Senescence
3.2. Mitochondria and Mitochondria-Related Phenomena
3.3. CNF1 on Immune Cells
3.4. CNF1 Effects on Different Cellular Pathways
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AKT | RAC- serine/threonine-protein kinase |
| AMPK | 5′-AMP-activated protein kinase catalytic subunit alpha-2 |
| AP-1 | activator protein 1 |
| Bcl-2 | apoptosis regulator Bcl-2 (B-cell lymphoma 2) |
| Bcl-XL | apoptosis regulator Bcl-X (B-cell lymphoma-extra-large) |
| C/EBPα | CCAAT/enhancer-binding protein alpha |
| cAMP | cyclic adenosine monophosphate |
| CD11a | integrin alpha-L |
| CD11b | integrin alpha-M |
| CD18 | integrin beta-2 |
| CD29 | integrin beta-1 |
| CD32 | low affinity immunoglobulin gamma Fc region receptor II-b |
| CD36 | platelet glycoprotein 4 |
| CD49d | integrin alpha-4 |
| CD83 | cluster of differentiation 83 antigen |
| CD86 | T-lymphocyte activation antigen CD86 |
| CD96 | T-cell surface protein tactile |
| c-Myc | proto-oncogene protein Myc |
| c-Jun | AP-1 transcription factor subunit |
| CR3 | complement receptor 3 C |
| DLK1 | protein delta homolog 1 |
| Drp1 | dynamin-1-like protein |
| Dsg3 | desmoglein-3 |
| EEA-1 | early endosome antigen 1 |
| EGFR | epidermal growth factor receptor |
| EIF4E | eukaryotic translation initiation factor 4E |
| EMT | epithelial–mesenchymal transition |
| ERK1/2, (p42-44, MAPK) | mitogen-activated protein kinases 3 and 1 |
| FAK | focal adhesion kinase |
| FoxG1 | forkhead box protein G1 |
| GFAP | glial fibrillary acidic protein |
| HIF-1α | hypoxia-inducible factor 1-alpha |
| HLA-DR | HLA class II histocompatibility antigen gamma chain |
| HMGA-2 | high mobility group protein HMGI-C |
| HSF1 | heat shock factor protein 1 |
| HSP90α | heat shock protein HSP 90-alpha |
| ICAM-1 | intercellular adhesion molecule 1 |
| IFN-γ | interferon gamma |
| IL-1β | interleukin 1 beta |
| IL-2R | interleukin 2 receptor |
| IL-6 | interleukin 6 |
| IL-8 | interleukin 8 |
| IκB-α, IKK | NF-κB inhibitor alpha |
| JAM-1 | junctional adhesion molecule A |
| JNK | mitogen-activated protein kinase 8 |
| LAMP1 | lysosome-associated membrane glycoprotein 1 |
| LC3-II | microtubule-associated proteins 1A/1B light chain 3B |
| LXRβ | liver X receptor beta |
| MAL | myocardin-related transcription factor A |
| MCP-1 | monocyte chemoattractant protein 1 |
| Met72 | melanoma cell-surface 72 Kd-glycoprotein |
| MHC | major histocompatibility complex |
| MIP-3α | macrophage inflammatory protein-3 alpha |
| MMP-2 | matrix metalloproteinase-2 |
| MMP-9 | matrix metalloproteinase-9 |
| moDC | monocyte-derived dendritic cells |
| MORs | mu-opioid receptors |
| mTOR | mammalian target of rapamycin |
| mTORC1 | mTOR complex 1 |
| MyoD | myoblast determination protein 1 |
| NADH | nicotinamide adenine dinucleotide, reduced form |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NIS | sodium/iodide cotransporter |
| Notch1 | neurogenic locus notch homolog protein 1 |
| p16 | cyclin-dependent kinase inhibitor 2A |
| p21 | cyclin-dependent kinase inhibitor 1 |
| p38 | mitogen-activated protein kinase 11 |
| p53 | cellular tumor antigen p53 |
| PAK1 | serine/threonine-protein kinase PAK1 |
| pAKT | RAC- serine/threonine-protein kinase, phosphorylate form |
| PDGFR | platelet-derived growth factor receptor |
| PdtIns-4-P 5-kinase | phosphatidylinositol 4-phosphate 5-kinase |
| PKA | cAMP-dependent protein kinase |
| PMN | polymorphonuclear neutrophils |
| PPARγ | peroxisome proliferator-activated receptor gamma |
| pRb | retinoblastoma-associated protein |
| pRBC | Plasmodium falciparum-parasitized erythrocytes |
| Pref1 | preadipocyte factor |
| pS6K | ribosomal protein S6 kinase, phosphorylate form |
| pULK1 | unc-51 like autophagy activating kinase 1, phosphorylate form |
| PV-IgG | pemphigus vulgaris autoantibodies |
| Rab11 | Ras-related protein Rab-11 |
| RagC | Ras-related GTP-binding protein C |
| rpS6 | 40S ribosomal protein S6 |
| Rip1, Rip2 | receptor-interacting serine/threonine-protein kinase 1 and 2 |
| ROS | reactive oxygen species |
| SA-β-gal | senescence-associated beta-galactosidase |
| SDF-1α | stromal cell-derived factor 1 |
| SMG | simulated microgravity |
| Snail1 | snail family transcriptional repressor 1 |
| TF | transcription factor |
| Tg | thyroglobulin |
| ThOX | thyroid oxidase |
| TGF-β | transforming growth factor beta |
| TJ/AJ | tight junction/adherent junction |
| TNF-α | tumor necrosis factor alpha |
| Tom20 | mitochondrial import receptor subunit |
| TRAF1 | TNF receptor-associated factor 1 |
| UPP1 | uridine-phosphorylase 1 |
| VEGF | vascular endothelial growth factor |
| VASP | vasodilator-stimulated phosphoprotein |
| ZEB1 | zinc finger E-box-binding homeobox 1 |
| ZO-1 | Zonula occudens-1 (tight junction protein 1) |
References
- Boquet, P. The cytotoxic necrotizing factor 1 (CNF1) from Escherichia coli. Toxicon 2001, 39, 1673–1680. [Google Scholar] [CrossRef]
- Piteau, M.; Papatheodorou, P.; Schwan, C.; Schlosser, A.; Aktories, K.; Schmidt, G. Lu/BCAM Adhesion Glycoprotein Is a Receptor for Escherichia coli Cytotoxic Necrotizing Factor 1 (CNF1). PLoS Pathog. 2014, 10, 3884. [Google Scholar] [CrossRef]
- Chung, J.W.; Hong, S.J.; Kim, K.J.; Goti, D.; Stins, M.F.; Shin, S.; Dawson, V.L.; Dawson, T.M.; Kim, K.S. 37-kDa laminin receptor precursor modulates cytotoxic necrotizing factor 1-mediated RhoA activation and bacterial uptake. J. Biol. Chem. 2003, 278, 16857–16862. [Google Scholar] [CrossRef] [Green Version]
- Contamin, S.; Galmiche, A.; Doye, A.; Flatau, G.; Benmerah, A.; Boquet, P. The p21 Rho-activating toxin cytotoxic necrotizing factor 1 is endocytosed by a clathrin-independent mechanism and enters the cytosol by an acidic-dependent membrane translocation step. Mol. Biol. Cell 2000, 11, 1775–1787. [Google Scholar] [CrossRef] [Green Version]
- Flatau, G.; Lemichez, E.; Gauthier, M.; Chardin, P.; Paris, S.; Fiorentini, C.; Boquet, P. Toxin-induced activation of the G protein p21 Rho by deamidation of glutamine. Nature 1997, 387, 729–733. [Google Scholar] [CrossRef]
- Schmidt, G.; Sehr, P.; Wilm, M.; Selzer, J.; Mann, M.; Aktories, K. Gln 63 of Rho is deamidated by Escherichia coli cytotoxic necrotizing factor-1. Nature 1997, 387, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Lerm, M.; Selzer, J.; Hoffmeyer, A.; Rapp, U.R.; Aktories, K.; Schmidt, G. Deamidation of Cdc42 and Rac by Escherichia coli cytotoxic necrotizing factor 1: Activation of c-Jun N-terminal kinase in HeLa cells. Infect. Immun. 1999, 67, 496–503. [Google Scholar] [CrossRef] [Green Version]
- Doye, A.; Mettouchi, A.; Bossis, G.; Clément, R.; Buisson-Touati, C.; Flatau, G.; Gagnoux, L.; Piechaczyk, M.; Boquet, P.; Lemichez, E. CNF1 exploits the ubiquitin-proteasome machinery to restrict Rho GTPase activation for bacterial host cell invasion. Cell 2002, 111, 553–564. [Google Scholar] [CrossRef] [Green Version]
- Lerm, M.; Pop, M.; Fritz, G.; Aktories, K.; Schmidt, G. Proteasomal degradation of cytotoxic necrotizing factor 1-activated rac. Infect. Immun. 2002, 70, 4053–4058. [Google Scholar] [CrossRef] [Green Version]
- Munro, P.; Flatau, G.; Doye, A.; Boyer, L.; Oregioni, O.; Mege, J.-L.; Landraud, L.; Lemichez, E. Activation and Proteasomal Degradation of Rho GTPases by Cytotoxic Necrotizing Factor-1 Elicit a Controlled Inflammatory Response. J. Biol. Chem. 2004, 279, 35849–35857. [Google Scholar] [CrossRef] [Green Version]
- Boyer, L.; Turchi, L.; Desnues, B.; Doye, A.; Ponzio, G.; Mege, J.L.; Yamashita, M.; Zhang, Y.E.; Bertoglio, J.; Flatau, G.; et al. CNF1-induced ubiquitylation and proteasome destruction of activated RhoA is impaired in Smurf1−/− cells. Mol. Biol. Cell 2006, 17, 2489–2497. [Google Scholar] [CrossRef] [Green Version]
- Hodge, R.G.; Ridley, A.J. Regulating Rho GTPases and their regulators. Nat. Rev. Mol. Cell Biol. 2016, 17, 496–510. [Google Scholar] [CrossRef] [PubMed]
- Ridley, A. Rho GTPases and actin dynamics in membrane protrusions and vesicle trafficking. Trends Cell Biol. 2006, 16, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Duquette, P.; Lamarche-Vane, N. Rho GTPases in embryonic development. Small GTPases 2014, 5, e972857. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; Sun, Z.; Li, H.; Feng, D. Rho GTPase-activating proteins: Regulators of Rho GTPase activity in neuronal development and CNS diseases. Mol. Cell. Neurosci. 2017, 80, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Niftullayev, S.; Lamarche-Vane, N. Regulators of rho GTPases in the nervous system: Molecular implication in axon guidance and neurological disorders. Int. J. Mol. Sci. 2019, 20, 1497. [Google Scholar] [CrossRef] [Green Version]
- Chai, L.; Cao, C.; Bi, S.; Dai, X.; Gan, L.; Guo, R.; Li, R. Small Rho GTPase Rac1 determines human epidermal stem cell fate in vitro. Int. J. Mol. Med. 2010, 25, 723–727. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wang, L.; Geiger, H.; Cancelas, J.; Mo, J.; Zheng, Y. Rho GTPase Cdc42 coordinates hematopoietic stem cell quiescence and niche interaction in the bone marrow. Proc. Natl. Acad. Sci. USA 2007, 104, 5091–5096. [Google Scholar] [CrossRef] [Green Version]
- Bros, M.; Haas, K.; Moll, L.; Grabbe, S. RhoA as a Key Regulator of Innate and Adaptive Immunity. Cells 2019, 8, 733. [Google Scholar] [CrossRef] [Green Version]
- Etienne-Manneville, S.; Hall, A. Rho GTPases in cell biology. Nature 2002, 420, 629–635. [Google Scholar] [CrossRef]
- Popoff, M.R. Bacterial factors exploit eukaryotic Rho GTPase signaling cascades to promote invasion and proliferation within their host. Small GTPases 2014, 5, e983863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabbri, A.; Travaglione, S.; Rosadi, F.; Ballan, G.; Maroccia, Z.; Giambenedetti, M.; Guidotti, M.; Ødum, N.; Krejsgaard, T.; Fiorentini, C. The Escherichia coli protein toxin cytotoxic necrotizing factor 1 induces epithelial mesenchymal transition. Cell Microbiol. 2019, 22, e13138. [Google Scholar] [CrossRef]
- Buc, E.; Dubois, D.; Sauvanet, P.; Raisch, J.; Delmas, J.; Darfeuille-Michaud, A.; Pezet, D.; Bonnet, R. High Prevalence of Mucosa-Associated, E. coli Producing Cyclomodulin and Genotoxin in Colon Cancer. PLoS ONE 2013, 8, e56964. [Google Scholar] [CrossRef] [Green Version]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement David Moher and colleagues introduce PRISMA, an update of the QUOROM guidelines for reporting systematic reviews and meta-analyses. BMJ 2009, 339, b2535. [Google Scholar] [CrossRef] [Green Version]
- Falzano, L.; Quaranta, M.G.; Travaglione, S.; Filippini, P.; Fabbri, A.; Viora, M.; Donelli, G.; Fiorentini, C. Cytotoxic necrotizing factor 1 enhances reactive oxygen species-dependent transcription and secretion of proinflammatory cytokines in human uroepithelial cells. Infect. Immun. 2003, 71, 4178–4181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falzano, L.; Filippini, P.; Travaglione, S.; Miraglia, A.G.; Fabbri, A.; Fiorentini, C. Escherichia coli cytotoxic necrotizing factor 1 blocks cell cycle G2/M transition in uroepithelial cells. Infect. Immun. 2006, 74, 3765–3772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, M.; Meysick, K.C.; O’Brien, A.D. Cytotoxic necrotizing factor type 1 of uropathogenic Escherichia coli kills cultured human uroepithelial 5637 cells by an apoptotic mechanism. Infect. Immun. 2000, 68, 5869–5880. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Wang, J.; Zhou, K.; Lv, J.; Wang, L.; Gao, S.; Keller, E.T.; Zhang, Z.S.; Wang, Q.; Yao, Z. Cytotoxic necrotizing factor 1 promotes bladder cancer angiogenesis through activating RhoC. FASEB J. 2020, 34, 7927–7940. [Google Scholar] [CrossRef] [Green Version]
- Travaglione, S.; Loizzo, S.; Vona, R.; Ballan, G.; Rivabene, R.; Giordani, D.; Guidotti, M.; Dupuis, M.L.; Maroccia, Z.; Baiula, M.; et al. The Bacterial Toxin CNF1 Protects Human Neuroblastoma SH-SY5Y Cells against 6-Hydroxydopamine-Induced Cell Damage: The Hypothesis of CNF1-Promoted Autophagy as an Antioxidant Strategy. Int. J. Mol. Sci. 2020, 21, 3390. [Google Scholar] [CrossRef]
- Pavone, F.; Luvisetto, S.; Marinelli, S.; Straface, E.; Fabbri, A.; Falzano, L.; Fiorentini, C.; Malorni, W. The Rac GTPase-activating bacterial protein toxin CNF1 induces analgesia up-regulating mu-opioid receptors. Pain 2009, 145, 219–229. [Google Scholar] [CrossRef]
- Vannini, E.; Olimpico, F.; Middei, S.; Ammassari-Teule, M.; de Graaf, E.L.; McDonnell, L.; Schmidt, G.; Fabbri, A.; Fiorentini, C.; Baroncelli, L.; et al. Electrophysiology of glioma: A Rho GTPase-activating protein reduces tumor growth and spares neuron structure and function. Neuro. Oncol. 2016, 18, 1634–1643. [Google Scholar] [CrossRef] [Green Version]
- Vannini, E.; Panighini, A.; Cerri, C.; Fabbri, A.; Lisi, S.; Pracucci, E.; Benedetto, N.; Vannozzi, R.; Fiorentini, C.; Caleo, M.; et al. The bacterial protein toxin, cytotoxic necrotizing factor 1 (CNF1) provides long-term survival in a murine glioma model. BMC Cancer 2014, 14, 449. [Google Scholar] [CrossRef] [PubMed]
- Stoll, T.; Markwirth, G.; Reipschläger, S.; Schmidt, G. A new member of a growing toxin family--Escherichia coli cytotoxic necrotizing factor 3 (CNF3). Toxicon 2009, 54, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, S.; May, M.; Schmidt, G.; Genth, H. Inhibition of cytokinesis by Clostridium difficile toxin B and cytotoxic necrotizing factors--reinforcing the critical role of RhoA in cytokinesis. Cell Motil. Cytoskeleton 2009, 66, 967–975. [Google Scholar] [CrossRef]
- Dong, N.; Liu, L.; Shao, F. A bacterial effector targets host DH-PH domain RhoGEFs and antagonizes macrophage phagocytosis. EMBO J. 2010, 29, 1363–1376. [Google Scholar] [CrossRef] [PubMed]
- May, M.; Kolbe, T.; Wang, T.; Schmidt, G.; Genth, H. Increased Cell-Matrix Adhesion upon Constitutive Activation of Rho Proteins by Cytotoxic Necrotizing Factors from E. Coli and Y. Pseudotuberculosis. J. Signal Transduct. 2012, 2012, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, S.C.; Roggenkamp, D.; May, M.; Huelsenbeck, J.; Brakebusch, C.; Rottner, K.; Ladwein, M.; Just, I.; Fritz, G.; Schmidt, G.; et al. Expression and cytoprotective activity of the small GTPase RhoB induced by the Escherichia coli cytotoxic necrotizing factor 1. Int. J. Biochem. Cell Biol. 2013, 45, 1767–1775. [Google Scholar] [CrossRef]
- Pfaumann, V.; Lang, A.E.; Schwan, C.; Schmidt, G.; Aktories, K. The actin and Rho-modifying toxins PTC3 and PTC5 of Photorhabdus luminescens: Enzyme characterization and induction of MAL/SRF-dependent transcription. Cell. Microbiol. 2015, 17, 579–594. [Google Scholar] [CrossRef]
- Gerhard, R.; Schmidt, G.; Hofmann, F.; Aktories, K. Activation of Rho GTPases by Escherichia coli cytotoxic necrotizing factor 1 increases intestinal permeability in Caco-2 cells. Infect. Immun. 1998, 66, 5125–5131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlegel, N.; Meir, M.; Spindler, V.; Germer, J.; Wascher, J. Differential role of Rho GTPases in intestinal epithelial barrier regulation in vitro. J. Cell. Physiol. 2011, 226, 1196–1203. [Google Scholar] [CrossRef]
- Hofman, P.; Flatau, G.; Selva, E.; Gauthier, M.; Le Negrate, G.; Fiorentini, C.; Rossi, B.; Boquet, P. Escherichia coli cytotoxic necrotizing factor 1 effaces microvilli and decreases transmigration of polymorphonuclear leukocytes in intestinal T84 epithelial cell monolayers. Infect. Immun. 1998, 66, 2494–2500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, A.; Walsh, S.; Verkade, P.; Boquet, P.; Nusrat, A. Constitutive activation of Rho proteins by CNF-1 influences tight junction structure and epithelial barrier function. J. Cell Sci. 2003, 116, 725–742. [Google Scholar] [CrossRef] [Green Version]
- Brest, P.; Turchi, L.; Le’Negrate, G.; Berto, F.; Moreilhon, C.; Mari, B.; Ponzio, G.; Hofman, P. Escherichia coli cytotoxic necrotizing factor 1 inhibits intestinal epithelial wound healing in vitro after mechanical injury. Infect. Immun. 2004, 72, 5733–5740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brest, P.; Mograbi, B.; Hofman, V.; Loubat, A.; Rossi, B.; Auberger, P.; Hofman, P. Rho GTPase is activated by cytotoxic necrotizing factor 1 in peripheral blood T lymphocytes: Potential cytotoxicity for intestinal epithelial cells. Infect. Immun. 2003, 71, 1161–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Aung, K.M.; Uhlin, B.E.; Wai, S.N. Reversible senescence of human colon cancer cells after blockage of mitosis/cytokinesis caused by the CNF1 cyclomodulin from Escherichia coli. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Falzano, L.; Fiorentini, C.; Donelli, G.; Michel, E.; Kocks, C.; Cossart, P.; Cabanié, L.; Oswald, E.; Boquet, P. Induction of phagocytic behaviour in human epithelial cells by Escherichia coli cytotoxic necrotizing factor type 1. Mol. Microbiol. 1993, 9, 1247–1254. [Google Scholar] [CrossRef]
- Fiorentini, C.; Donelli, G.; Matarrese, P.; Fabbri, A.; Paradisi, S.; Boquet, P. Escherichia coli cytotoxic necrotizing factor 1: Evidence for induction of actin assembly by constitutive activation of the p21 Rho GTPase. Infect. Immun. 1995, 63, 3936–3944. [Google Scholar] [CrossRef] [Green Version]
- Fiorentini, C.; Fabbri, A.; Flatau, G.; Donelli, G.; Matarrese, P.; Lemichez, E.; Falzano, L.; Boquet, P. Escherichia coli cytotoxic necrotizing factor 1 (CNF1), a toxin that activates the Rho GTPase. J. Biol. Chem. 1997, 272, 19532–19537. [Google Scholar] [CrossRef] [Green Version]
- Fiorentini, C.; Matarrese, P.; Straface, E.; Falzano, L.; Donelli, G.; Boquet, P.; Malorni, W. Rho-dependent cell spreading activated by E.coli cytotoxic necrotizing factor 1 hinders apoptosis in epithelial cells. Cell Death Differ. 1998, 5, 921–929. [Google Scholar] [CrossRef] [Green Version]
- Falzano, L.; Rivabene, R.; Santini, M.T.; Fabbri, A.; Fiorentini, C. An Escherichia coli cytotoxin increases superoxide anion generation via Rac in epithelial cells. Biochem. Biophys. Res. Commun. 2001, 283, 1026–1030. [Google Scholar] [CrossRef]
- Fiorentini, C.; Falzano, L.; Fabbri, A.; Stringaro, A.; Logozzi, M.; Travaglione, S.; Contamin, S.; Arancia, G.; Malorni, W.; Fais, S. Activation of Rho GTPases by Cytotoxic Necrotizing Factor 1 Induces Macropinocytosis and Scavenging Activity in Epithelial Cells. Mol. Biol. Cell 2001, 12, 2061–2073. [Google Scholar] [CrossRef] [Green Version]
- Falzano, L.; Rivabene, R.; Fabbri, A.; Fiorentini, C. Epithelial cells challenged with a Rac-activating E. coli cytotoxin acquire features of professional phagocytes. Toxicol. Vitr. 2002, 16, 421–425. [Google Scholar] [CrossRef]
- Boyer, L.; Travaglione, S.; Falzano, L.; Gauthier, N.C.; Popoff, M.R.; Lemichez, E.; Fiorentini, C.; Fabbri, A. Rac GTPase Instructs Nuclear Factor-κB Activation by Conveying the SCF Complex and IkBα to the Ruffling Membranes. Mol. Biol. Cell 2004, 15, 1124–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malorni, W.; Fiorentini, C. Is the Rac GTPase-activating toxin CNF1 a smart hijacker of host cell fate? FASEB J. 2006, 20, 606–609. [Google Scholar] [CrossRef] [PubMed]
- Giamboi-Miraglia, A.; Travaglione, S.; Filippini, P.; Fabbri, A.; Fiorentini, C.; Falzano, L. A multinucleating Escherichia coli cytotoxin perturbs cell cycle in cultured epithelial cells. Toxicol. Vitro 2007, 21, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Miraglia, A.G.; Travaglione, S.; Meschini, S.; Falzano, L.; Matarrese, P.; Quaranta, M.G.; Viora, M.; Fiorentini, C.; Fabbri, A. Cytotoxic necrotizing factor 1 prevents apoptosis via the Akt/IkappaB kinase pathway: Role of nuclear factor-kappaB and Bcl-2. Mol. Biol. Cell 2007, 18, 2735–2744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabbri, A.; Cori, S.; Zanetti, C.; Guidotti, M.; Sargiacomo, M.; Loizzo, S.; Fiorentini, C. Cell-to-cell propagation of the bacterial toxin CNF1 via extracellular vesicles: Potential impact on the therapeutic use of the toxin. Toxins 2015, 7, 4610–4621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNichol, B.A.; Rasmussen, S.B.; Meysick, K.C.; O’Brien, A.D. A single amino acid substitution in the enzymatic domain of cytotoxic necrotizing factor type 1 of Escherichia coli alters the tissue culture phenotype to that of the dermonecrotic toxin of Bordetella spp. Mol. Microbiol. 2006, 60, 939–950. [Google Scholar] [CrossRef]
- Abouzahr-Rifai, S.; Hasmim, M.; Boukerche, H.; Hamelin, J.; Janji, B.; Jalil, A.; Kieda, C.; Mami-Chouaib, F.; Bertoglio, J.; Chouaib, S. Resistance of tumor cells to cytolytic T lymphocytes involves Rho-GTPases and focal adhesion kinase activation. J. Biol. Chem. 2008, 283, 31665–31672. [Google Scholar] [CrossRef] [Green Version]
- Capo, C.; Meconi, S.; Sanguedolce, M.V.; Bardin, N.; Flatau, G.; Boquet, P.; Mege, J.L. Effect of cytotoxic necrotizing factor-1 on actin cytoskeleton in human monocytes: Role in the regulation of integrin-dependent phagocytosis. J. Immunol. 1998, 161, 4301–4308. [Google Scholar]
- Yang, H.; Li, Q.; Wang, C.; Wang, J.; Lv, J.; Wang, L.; Zhang, Z.S.; Yao, Z.; Wang, Q. Cytotoxic Necrotizing Factor 1 Downregulates CD36 Transcription in Macrophages to Induce Inflammation During Acute Urinary Tract Infections. Front. Immunol. 2018, 9, 1987. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, Z.; Wei, H.; Wang, J.; Lv, J.; Zhang, K.; Keller, E.T.; Yao, Z.; Wang, Q. Cytotoxic necrotizing factor 1 promotes prostate cancer progression through activating the Cdc42-PAK1 axis. J. Pathol. 2017, 243, 208–219. [Google Scholar] [CrossRef] [Green Version]
- Augspach, A.; List, J.; Wolf, P.; Bielek, H.; Schwan, C.; Elsässer-Beile, U.; Aktories, K.; Schmidt, G. Activation of RhoA,B,C by Yersinia Cytotoxic Necrotizing Factor (CNFy) induces apoptosis in LNCaP prostate cancer cells. Toxins 2013, 5, 2241–2257. [Google Scholar] [CrossRef]
- Tan, X.; Xu, A.; Zhao, T.; Zhao, Q.; Zhang, J.; Fan, C.; Deng, Y.; Freywald, A.; Genth, H.; Xiang, J. Simulated microgravity inhibits cell focal adhesions leading to reduced melanoma cell proliferation and metastasis via FAK/RhoA-regulated mTORC1 and AMPK pathways. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, T.; Li, R.; Tan, X.; Zhang, J.; Fan, C.; Zhao, Q.; Deng, Y.; Xu, A.; Lukong, K.E.; Genth, H.; et al. Simulated microgravity reduces focal adhesions and alters cytoskeleton and nuclear positioning leading to enhanced apoptosis via suppressing FAK/Rhoa-mediated mTORC1/NF-κB and ERK1/2 pathways. Int. J. Mol. Sci. 2018, 19, 1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buscà, R.; Bertolotto, C.; Abbe, P.; Englaro, W.; Ishizaki, T.; Narumiya, S.; Boquet, P.; Ortonne, J.P.; Ballotti, R. Inhibition of Rho is required for cAMP-induced melanoma cell differentiation. Mol. Biol. Cell 1998, 9, 1367–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stray, A.; Janning, A.; Barth, H.; Gerke, V. Endothelial Rho signaling is required for monocyte transendothelial migration. FEBS Lett. 2002, 517, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Boyer, L.; Magoc, L.; Dejardin, S.; Cappillino, M.; Paquette, N.; Hinault, C.; Charriere, G.; Ip, W.; Fracchia, S.; Hennessy, E.; et al. Pathogen-derived effectors trigger protective immunity via activation of the Rac2 enzyme and the IMD or Rip kinase signaling pathway. Immunity 2011, 35, 536–549. [Google Scholar] [CrossRef] [Green Version]
- Messina, V.; Loizzo, S.; Travaglione, S.; Bertuccini, L.; Condello, M.; Superti, F.; Guidotti, M.; Alano, P.; Silvestrini, F.; Fiorentini, C. The bacterial protein CNF1 as a new strategy against Plasmodium falciparum cytoadherence. PLoS ONE 2019, 14, e0213529. [Google Scholar] [CrossRef]
- Baumer, Y.; Burger, S.; Curry, F.E.; Golenhofen, N.; Drenckhahn, D.; Waschke, J. Differential role of Rho GTPases in endothelial barrier regulation dependent on endothelial cell origin. Histochem. Cell Biol. 2008, 129, 179–191. [Google Scholar] [CrossRef]
- Schlegel, N.; Burger, S.; Golenhofen, N.; Walter, U.; Drenckhahn, D.; Waschke, J. The role of VASP in regulation of cAMP- and Rac 1-mediated endothelial barrier stabilization. Am. J. Physiol. Cell Physiol. 2008, 294, C178–C188. [Google Scholar] [CrossRef]
- Waschke, J.; Burger, S.; Curry, F.R.E.; Drenckhahn, D.; Adamson, R.H. Activation of Rac-1 and Cdc42 stabilizes the microvascular endothelial barrier. Histochem. Cell Biol. 2006, 125, 397–406. [Google Scholar] [CrossRef]
- Gliem, M.; Heupel, W.M.; Spindler, V.; Harms, G.S.; Waschke, J. Actin reorganization contributes to loss of cell adhesion in pemphigus vulgaris. Am. J. Physiol. Cell Physiol. 2010, 299, C606–C613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiariello, M.; Marinissen, M.J.; Gutkind, J.S. Regulation of c-myc expression by PDGF through Rho GTPases. Nat. Cell Biol. 2001, 3, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Bannai, Y.; Aminova, L.R.; Faulkner, M.J.; Ho, M.; Wilson, B.A. Rho/ROCK-dependent inhibition of 3T3-L1 adipogenesis by G-protein-deamidating dermonecrotic toxins: Differential regulation of Notch1, Pref1/Dlk1, and β-catenin signaling. Front. Cell. Infect. Microbiol. 2012, 2, 80. [Google Scholar] [CrossRef] [Green Version]
- Travaglione, S.; Messina, G.; Fabbri, A.; Falzano, L.; Giammarioli, A.M.; Grossi, M.; Rufini, S.; Fiorentini, C. Cytotoxic necrotizing factor 1 hinders skeletal muscle differentiation in vitro by perturbing the activation/deactivation balance of Rho GTPases. Cell Death Differ. 2005, 12, 78–86. [Google Scholar] [CrossRef]
- Kazmierczak, B.I.; Jou, T.S.; Mostov, K.; Engel, J.N. Rho GTPase activity modulates Pseudomonas aeruginosa internalization by epithelial cells. Cell. Microbiol. 2001, 3, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Moreau, V.; Tatin, F.; Varon, C.; Génot, E. Actin Can Reorganize into Podosomes in Aortic Endothelial Cells, a Process Controlled by Cdc42 and RhoA. Mol. Cell. Biol. 2003, 23, 6809–6822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumer, Y.; Drenckhahn, D.; Waschke, J. cAMP induced Rac 1-mediated cytoskeletal reorganization in microvascular endothelium. Histochem. Cell Biol. 2008, 129, 765–778. [Google Scholar] [CrossRef]
- Vouret-Craviari, V.; Grall, D.; Flatau, G.; Pouysségur, J.; Boquet, P.; Van Obberghen-Schilling, E. Effects of cytotoxic necrotizing factor 1 and lethal toxin on actin cytoskeleton and VE-cadherin localization in human endothelial cell monolayers. Infect. Immun. 1999, 67, 3002–3008. [Google Scholar] [CrossRef] [Green Version]
- Travaglione, S.; Loizzo, S.; Rizza, T.; Del Brocco, A.; Ballan, G.; Guidotti, M.; Vona, R.; Di Nottia, M.; Torraco, A.; Carrozzo, R.; et al. Enhancement of mitochondrial ATP production by the Escherichia coli cytotoxic necrotizing factor 1. FEBS J. 2014, 281, 3473–3488. [Google Scholar] [CrossRef]
- Malorni, W.; Quaranta, M.G.; Straface, E.; Falzano, L.; Fabbri, A.; Viora, M.; Fiorentini, C. The Rac-activating toxin cytotoxic necrotizing factor 1 oversees NK cell-mediated activity by regulating the actin/microtubule interplay. J. Immunol. 2003, 171, 4195–4202. [Google Scholar] [CrossRef] [PubMed]
- Gall-Mas, L.; Fabbri, A.; Namini, M.R.J.; Givskov, M.; Fiorentini, C.; Krejsgaard, T. The bacterial toxin CNF1 induces activation and maturation of human monocyte-derived dendritic cells. Int. J. Mol. Sci. 2018, 19, 1408. [Google Scholar] [CrossRef] [Green Version]
- Fabbri, A.; Travaglione, S.; Maroccia, Z.; Guidotti, M.; Pierri, C.L.; Primiano, G.; Servidei, S.; Loizzo, S.; Fiorentini, C. The bacterial protein CNF1 as a potential therapeutic strategy against mitochondrial diseases: A pilot study. Int. J. Mol. Sci. 2018, 19, 1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufies, O.; Doye, A.; Courjon, J.; Torre, C.; Michel, G.; Loubatier, C.; Jacquel, A.; Chaintreuil, P.; Majoor, A.; Guinamard, R.R.; et al. Escherichia coli Rho GTPase-activating toxin CNF1 mediates NLRP3 inflammasome activation via p21-activated kinases-1/2 during bacteraemia in mice. Nat. Microbiol. 2021, 6, 401–412. [Google Scholar] [CrossRef]
- Hahn, A.; Barth, H.; Kress, M.; Mertens, P.R.; Goppelt-Struebe, M. Role of Rac and Cdc42 in lysophosphatidic acid-mediated cyclo-oxygenase-2 gene expression. Biochem. J. 2002, 362, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Malchiodi-Albedi, F.; Paradisi, S.; Di Nottia, M.; Simone, D.; Travaglione, S.; Falzano, L.; Guidotti, M.; Frank, C.; Cutarelli, A.; Fabbri, A.; et al. CNF1 improves astrocytic ability to support neuronal growth and differentiation in vitro. PLoS ONE 2012, 7, e34115. [Google Scholar] [CrossRef]
- Boutillier, S.; Rapp, J.; Staeb, T.; Olenik, C.; Schmidt, G.; Meyer, D.K.; Leemhuis, J. Cytotoxic necrotizing factor-2 of Escherichia coli alters the morphology of cultured hippocampal neurons. Naunyn. Schmiedebergs. Arch. Pharmacol. 2003, 368, 513–519. [Google Scholar] [CrossRef]
- Musilli, M.; Ciotti, M.T.; Pieri, M.; Martino, A.; Borrelli, S.; Dinallo, V.; Diana, G. Therapeutic effects of the Rho GTPase modulator CNF1 in a model of Parkinson’s disease. Neuropharmacology 2016, 109, 357–365. [Google Scholar] [CrossRef]
- Czopka, T.; Von Holst, A.; Schmidt, G.; Ffrench-Constant, C.; Faissner, A. Tenascin C and tenascin R similarly prevent the formation of myelin membranes in a RhoA-dependent manner, but antagonistically regulate the expression of myelin basic protein via a separate pathway. Glia 2009, 57, 1790–1801. [Google Scholar] [CrossRef]
- Fortemaison, N.; Blancquaert, S.; Dumont, J.E.; Maenhaut, C.; Aktories, K.; Roger, P.P.; Dremier, S. Differential involvement of the actin cytoskeleton in differentiation and mitogenesis of thyroid cells: Inactivation of Rho proteins contributes to cyclic adenosine monophosphate-dependent gene expression but prevents mitogenesis. Endocrinology 2005, 146, 5485–5495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doye, A.; Boyer, L.; Mettouchi, A.; Lemichez, E. Ubiquitin-mediated proteasomal degradation of Rho proteins by the CNF1 toxin. Methods Enzymol. 2006, 406, 447–456. [Google Scholar] [CrossRef]
- Haga, R.B.; Ridley, A.J. Rho GTPases: Regulation and roles in cancer cell biology. Small GTPases 2016, 7, 207–221. [Google Scholar] [CrossRef] [Green Version]
- Ding, F.; Yin, Z.; Wang, H.-R. Ubiquitination in Rho Signaling. Curr. Top. Med. Chem. 2011, 11, 2879–2887. [Google Scholar] [CrossRef]
- Yang, S.; Rosenwald, A. Small GTPase proteins in macroautophagy. Small GTPases 2018, 9, 409–414. [Google Scholar] [CrossRef]
- Fiorentini, C.; Matarrese, P.; Straface, E.; Falzano, L.; Fabbri, A.; Donelli, G.; Cossarizza, A.; Boquet, P.; Malorni, W. Toxin-Induced Activation of Rho GTP-Binding Protein Increases Bcl-2 Expression and Influences Mitochondrial Homeostasis. Exp. Cell Res. 1998, 242, 341–350. [Google Scholar] [CrossRef]
- Horiguchi, Y. Escherichia coli cytotoxic necrotizing factors and Bordetella dermonecrotic toxin: The dermonecrosis-inducing toxins activating Rho small GTPases. Toxicon 2001, 39, 1619–1627. [Google Scholar] [CrossRef]
- Chircop, M. Rho GTPases as regulators of mitosis and cytokinesis in mammalian cells. Small GTPases 2014, 5, e29770. [Google Scholar] [CrossRef] [Green Version]
- El-Aouar Filho, R.A.; Nicolas, A.; De Paula Castro, T.L.; Deplanche, M.; De Carvalho Azevedo, V.A.; Goossens, P.L.; Taieb, F.; Lina, G.; Le Loir, Y.; Berkova, N. Heterogeneous family of cyclomodulins: Smart weapons that allow bacteria to hijack the eukaryotic cell cycle and promote infections. Front. Cell. Infect. Microbiol. 2017, 7, 208. [Google Scholar] [CrossRef] [Green Version]
- Island, M.D.; Cui, X.; Warren, J.W. Effect of Escherichia coli cytotoxic necrotizing factor 1 on repair of human bladder cell monolayers in vitro. Infect. Immun. 1999, 67, 3657–3661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munro, P.; Flatau, G.; Lemichez, E. Intranasal immunization with tetanus toxoid and CNF1 as a new mucosal adjuvant protects BALB/c mice against lethal challenge. Vaccine 2007, 25, 8702–8706. [Google Scholar] [CrossRef] [PubMed]
- Diabate, M.; Munro, P.; Garcia, E.; Jacquel, A.; Michel, G.; Obba, S.; Goncalves, D.; Luci, C.; Marchetti, S.; Demon, D.; et al. Escherichia coli α-Hemolysin Counteracts the Anti-Virulence Innate Immune Response Triggered by the Rho GTPase Activating Toxin CNF1 during Bacteremia. PLoS Pathog. 2015, 11, 1–20. [Google Scholar] [CrossRef]
- Ahmed, A.U.; Williams, B.R.G.; Hannigan, G.E. Transcriptional activation of inflammatory genes: Mechanistic insight into selectivity and diversity. Biomolecules 2015, 5, 3087–3111. [Google Scholar] [CrossRef] [PubMed]
- Collins, P.W.; Noble, K.E.; Reittie, J.R.; Hoffbrand, A.V.; Pasi, K.J.; Yong, K.L. Induction of tissue factor expression in human monocyte/endothelium cocultures. Br. J. Haematol. 1995, 91, 963–970. [Google Scholar] [CrossRef]
- Fabbri, A.; Travaglione, S.; Fiorentini, C. Escherichia coli cytotoxic necrotizing factor 1 (CNF1): Toxin biology, in vivo applications and therapeutic potential. Toxins 2010, 2, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Omar, A.; Jovanovic, K.; Da Costa Dias, B.; Gonsalves, D.; Moodley, K.; Caveney, R.; Mbazima, V.; Weiss, S.F. Patented biological approaches for the therapeutic modulation of the 37 kDa/67 kDa laminin receptor. Expert Opin. Ther. Pat. 2011, 21, 35–53. [Google Scholar] [CrossRef]
- Gopalakrishna, R.; Bhat, N.R.; Zhou, S.; Mack, W.J. Cell signaling associated with internalization of 67 kDa laminin receptor (67LR) by soluble laminin and its implication for protection against neurodegenerative diseases. Neural Regen. Res. 2019, 14, 1513–1514. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, S.; Rimondini, R.; Travaglione, S.; Fabbri, A.; Guidotti, M.; Ferri, A.; Campana, G.; Fiorentini, C. CNF1 Increases Brain Energy Level, Counteracts Neuroinflammatory Markers and Rescues Cognitive Deficits in a Murine Model of Alzheimer’s Disease. PLoS ONE 2013, 8, e65898. [Google Scholar] [CrossRef]
- Cloutier, J.M.; Charville, G.W. Diagnostic classification of soft tissue malignancies: A review and update from a surgical pathology perspective. Curr. Probl. Cancer 2019, 43, 250–272. [Google Scholar] [CrossRef]
- Kashina, A.S. Regulation of actin isoforms in cellular and developmental processes. Semin. Cell Dev. Biol. 2020, 102, 113–121. [Google Scholar] [CrossRef]
- Travaglione, S.; Fabbri, A.; Fiorentini, C. The Rho-activating CNF1 toxin from pathogenic E. coli: A risk factor for human cancer development? Infect. Agent. Cancer 2008, 3, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabbri, A.; Travaglione, S.; Ballan, G.; Loizzo, S.; Fiorentini, C.; Fabbri, A.; Travaglione, S.; Ballan, G.; Loizzo, S.; Fiorentini, C. The Cytotoxic Necrotizing Factor 1 from E. Coli: A Janus Toxin Playing with Cancer Regulators. Toxins 2013, 5, 1462–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piciocchi, A.; Germinario, E.A.P.; Garcia Etxebarria, K.; Rossi, S.; Sanchez-Mete, L.; Porowska, B.; Stigliano, V.; Trentino, P.; Oddi, A.; Accarpio, F.; et al. Association of polygenic risk score and bacterial toxins at screening colonoscopy with colorectal cancer progression: A multicenter case-control study. Toxins 2021, 13, 569. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cell Line | Tissue Origin and Morphology | References | CNF1 Described Effects | CNF1 Preparation |
|---|---|---|---|---|
| T24 | hu bladder, carcinoma, epithelial | [11,25,26] * [27,28] § |
| recombinant, * purified by chromatography § His-tagged protein |
| UMUC3 | hu bladder, carcinoma, epithelial | [28] | MMP-9 activity increase | recombinant, His-tagged protein |
| 5637 | hu bladder, carcinoma, epithelial | [26] * [27,28] § |
| recombinant, * purified by chromatography § His-tagged protein |
| J82 | hu bladder, carcinoma, epithelial | [27] |
| recombinant, His-tagged protein |
| SH-SY5Y | hu brain, neuroblastoma, epithelial | [29] |
| recombinant, purified by chromatography |
| SK-N-SH | hu brain, neuroblastoma, epithelial | [30] |
| recombinant, purified by chromatography |
| U87 | hu brain, glioblastoma like, epithelial | [31] |
| recombinant, purified by chromatography |
| GBM (Glioblastoma) | hu brain, glioblastoma multiforme | [31,32] |
| recombinant, purified by chromatography |
| MCF7 | hu breast, ductal carcinoma, epithelial | [11] | Rho, Rac, and Cdc42 efficient depletion | recombinant, purified by chromatography |
| HeLa | hu cervix, adenocarcinoma, epithelial | [7,9,33,34,35,36,37,38] |
| recombinant, GST fusion protein |
| Caco-2 | hu colon, adenocarcinoma, epithelial | [33,39,40] |
| recombinant, GST fusion protein |
| SW480 | hu colon, adenocarcinoma, epithelial | [22] | enhanced migration and invasion | recombinant, purified by chromatography |
| SW620 | hu colon, adenocarcinoma, epithelial | [11] | Rho, Rac, and Cdc42 efficient depletion | recombinant, purified by chromatography |
| HT-29 | hu colon, adenocarcinoma, epithelial | [22] * [37] ‡ |
| recombinant: * purified by chromatography ‡ GST fusion protein |
| T84 | hu colon, adenocarcinoma, epithelial | [41,42,43,44] |
| recombinant, purified by chromatography |
| HCT-116 | hu colon carcinoma, epithelial | [45] |
| recombinant, His-tagged protein |
| ACHN | hu kidney, adenocarcinoma, epithelial | [27] |
| recombinant, His-tagged protein |
| A-498 | hu kidney, carcinoma, epithelial | [27,28] |
| recombinant, His-tagged protein |
| HEp-2 (Human Epidermoid carcinoma #2) | hu larynx, carcinoma, epithelial | [2,7,10,21,26,45,46,47,48,49,50,51,52,53,54,55,56,57,58] |
| recombinant, purified by chromatography |
| IGR-Heu | hu non-small-cell, lung carcinoma, epithelial | [59] |
| recombinant, GST fusion protein |
| IGR-Heu R8 | hu non-small-cell, lung carcinoma, epithelial | [59] |
| recombinant, GST fusion protein |
| THP-1 | hu peripheral blood, acute monoblastic/monocytic leukemia, monocyte | [60] * [61] § |
| recombinant, *purified by chromatography § His-tagged protein |
| JURKAT | hu peripheral blood, acute T cell leukemia, lymphoblast | [44] |
| recombinant, purified by chromatography |
| PC3 | hu prostate, adenocarcinoma, epithelial | [62] * [63] ‡ |
| recombinant, *purified by chromatography ‡ GST fusion protein |
| LNCaP (Lymph Node Carcinoma of the Prostate) | hu prostate, carcinoma, epithelial | [62] * [63] ‡ |
| recombinant, *purified by chromatography ‡ GST fusion protein |
| 22Rv1 | hu prostate, carcinoma, epithelial | [62] |
| recombinant, purified by chromatography |
| VCaP (Vertebral Cancer of the Prostate) | hu prostate, carcinoma, epithelial | [62] | enhanced migration and invasion | recombinant, purified by chromatography |
| Me-665 | hu skin, melanoma, epithelial | [52] | formation of stress fibers, ruffles and filopodia | recombinant, purified by chromatography |
| RAW264.7 | mouse abelson murine leukemia virus-induced tumor, monocyte/macrophage | [61] |
| recombinant, His-tagged protein |
| Y-1 | mouse adrenal cortical carcinoma, epithelial | [27] |
| recombinant, His-tagged protein |
| GL261 (Glioma 261) | mouse brain, glioblastoma, fibroblastoid | [31,32] |
| recombinant, purified by chromatography |
| BL6-10 | mouse skin, melanoma, epithelial | [64,65] |
| recombinant, GST-fusion protein |
| B16-F10 | mouse skin, melanoma, mixture of spindle-shaped and epithelial-like | [66] |
| not specified |
| 804G | rat bladder, carcinoma, epithelial | [8,11] |
| recombinant, purified by chromatography |
| Cell Line | Tissue Origin and Morphology | References | CNF1 Described Effects | CNF1 Preparation |
|---|---|---|---|---|
| HMEC-1 (Human Microvascular Endothelial Cell line-1) | hu dermal endothelium, immortalized (SV40 T-antigen), endothelial-like | [67] |
| recombinant, GST fusion protein |
| HEK 293 (Human Embryonic Kidney 293) | hu embryonic kidney, transformed, tumorigenic, epithelial | [11] * [9] ‡ |
| recombinant, * purified by chromatography ‡ GST fusion protein |
| HEK 293T (Human Embryonic Kidney 293T) | hu embryonic kidney, transformed, HEK 293 derivative expressing SV40 T-antigen, epithelial | [68] |
| recombinant, His-tagged protein |
| HBMEC-60 (Human Bone Marrow Endothelial Cell line-60) | hu bone marrow, immortalized (HPV16 E6/E7), endothelial | [69] |
| recombinant, purified by chromatography |
| HBEC-5i (Human Brain Endothelial Cell line-5i) | hu brain cerebral cortex, immortalized (SV40 T-antigen), endothelial | [22,69] |
| recombinant, purified by chromatography |
| SV-HUC-1 | hu ureter, immortalized (SV40 T-antigen), epithelial | [27] | multinucleation | recombinant, His-tagged protein |
| MesEnd (Mesenteric Endothelial) | mouse mesenteric microvascular, immortalized (SV40 T-antigen), | [70] |
| recombinant, GST fusion protein |
| MyEnd (Myocardial Endothelial) | mouse microvascular myocardial, immortalized (SV40 T-antigen) | [70,71,72] |
| recombinant, GST fusion protein |
| HaCaT | hu skin, spontaneously immortalized, keratinocyte | [73] |
| not specified |
| NIH 3T3 | mouse embryonic, spontaneously immortalized, fibroblasts | [6,33,37,74] |
| recombinant, GST fusion protein |
| 3T3-Swiss albino | mouse embryo, spontaneously immortalized, fibroblasts | [53] |
| recombinant, His-tagged protein |
| 3T3-L1 | mouse embryo, substrain of 3T3-Swiss albino, preadipocytes fibroblasts | [75] |
| recombinant, His-tagged protein |
| C2C12 | mouse muscle, spontaneously immortalized, myoblast | [76] |
| recombinant, purified by chromatography |
| Vero | monkey kidney, spontaneously immortalized, epithelial | [5,11,56] |
| recombinant, purified by chromatography |
| MDCK (Madin-Darby Canine Kidney) | dog kidney, spontaneously immortalized, epithelial | [77] |
| recombinant, GST fusion protein |
| PAE (p23 clone) (Porcine Aorta-derived Endothelial) | pig aorta | [78] |
| recombinant, GST fusion protein |
| Cell line | Tissue Origin and Morphology | References | CNF1 Described Effects | CNF1 Preparation |
|---|---|---|---|---|
| HPECC (HPCEC) | hu colon, finite cell line, epithelial | [22] | cell motility decrease | recombinant, purified by chromatography |
| HDMEC (Human Dermal Microvascular Endothelial Cells) | hu dermal, finite cell line, microvascular endothelial cells | [70,79] |
| recombinant, GST fusion protein |
| HUVEC (Human Umbilical Vein Endothelial Cell) | hu umbilical, finite cell line, endothelial | [8,10,11,28,80] |
| recombinant, purified by chromatography, His-tagged protein |
| IEC-6 (Intestinal Epithelioid Cell line #6) | rat small intestine, finite cell line, epithelial | [22,81] |
| recombinant, purified by chromatography |
| T-lymphocytes | hu blood, primary, lymphocyte | [44] |
| recombinant, purified by chromatography |
| NK | hu blood, primary, large granular lymphocyte | [82] |
| recombinant, purified by chromatography |
| monocytes | hu blood, primary, monocyte | [60] |
| recombinant, purified by chromatography |
| macrophages | hu blood, primary, macrophage | [11] | Rho, Rac, and Cdc42 depletion | recombinant, purified by chromatography |
| DC (dendritic cell) monocytes | hu blood, primary, monocyte | [83] |
| recombinant, purified by chromatography |
| HBMEC (Human Brain Microvascular Endothelial Cells) | hu brain, primary, microvascular endothelium | [3] |
| recombinant, GST fusion protein |
| keratinocytes | hu neonatal foreskin, primary, keratinocyte | [11] | RhoA, Rac1, and Cdc42 depletion | recombinant, purified by chromatography |
| MERRF (Myoclonic Epilepsy with Ragged-Red Fibers) fibroblasts | hu skin, primary, from myoclonic epilepsy with ragged-red fibers, fibroblast | [84] |
| recombinant, purified by chromatography |
| fibroblasts | hu skin/neonatal foreskin, primary, fibroblast | [11,84] |
| recombinant, purified by chromatography |
| BMDM (Bone-Marrow-Derived Macrophages) | mouse bone marrow derived macrophages | [85] * [61] § |
| recombinant, * purified by chromatography § His-tagged protein |
| MEFs (Mouse Embryonic Fibroblasts) | mouse embryo, primary, fibroblast | [11] |
| recombinant, purified by chromatography |
| mouse peritoneal macrophages | mouse peritoneal lavage, primary, macrophage | [61] |
| recombinant, His-tagged protein |
| rat mesangial primary cells | rat kidney, primary | [86] | increase of Cox2 mRNA levels | recombinant, His-tagged protein |
| rat embryonic primary astrocytes | rat embryo cortex, primary, astrocytes | [87] |
| recombinant, purified by chromatography |
| rat embryonic primary neurons | rat embryonic hippocampus | [87] |
| recombinant, purified by chromatography |
| rat embryonic primary neurons | rat newborn hippocampus | [88] | dendrite and axon retraction | recombinant, GST-fusion |
| rat embryonic primary neurons | rat embryonic substantia nigra | [89] |
| recombinant, His-SUMO tag protein |
| OPC (Oligodendrocyte Precursor Cells) | rat/mouse newborn cortex, primary | [90] |
| recombinant, GST fusion protein |
| PAEC (Porcine Aorta Endothelial Cells) | pig pulmonary artery, primary, endothelial | [70] |
| recombinant, GST fusion protein |
| dog thyroid epithelial cells | dog thyroid, primary, epithelial-like | [91] |
| recombinant, GST fusion protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carlini, F.; Maroccia, Z.; Fiorentini, C.; Travaglione, S.; Fabbri, A. Effects of the Escherichia coli Bacterial Toxin Cytotoxic Necrotizing Factor 1 on Different Human and Animal Cells: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 12610. https://doi.org/10.3390/ijms222212610
Carlini F, Maroccia Z, Fiorentini C, Travaglione S, Fabbri A. Effects of the Escherichia coli Bacterial Toxin Cytotoxic Necrotizing Factor 1 on Different Human and Animal Cells: A Systematic Review. International Journal of Molecular Sciences. 2021; 22(22):12610. https://doi.org/10.3390/ijms222212610
Chicago/Turabian StyleCarlini, Francesca, Zaira Maroccia, Carla Fiorentini, Sara Travaglione, and Alessia Fabbri. 2021. "Effects of the Escherichia coli Bacterial Toxin Cytotoxic Necrotizing Factor 1 on Different Human and Animal Cells: A Systematic Review" International Journal of Molecular Sciences 22, no. 22: 12610. https://doi.org/10.3390/ijms222212610