Insecticide Exposure Triggers a Modulated Expression of ABC Transporter Genes in Larvae of Anopheles gambiae s.s.

Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquitoes
2.2. Bioassays
2.3. Induction of ABC Transporter Gene Expression
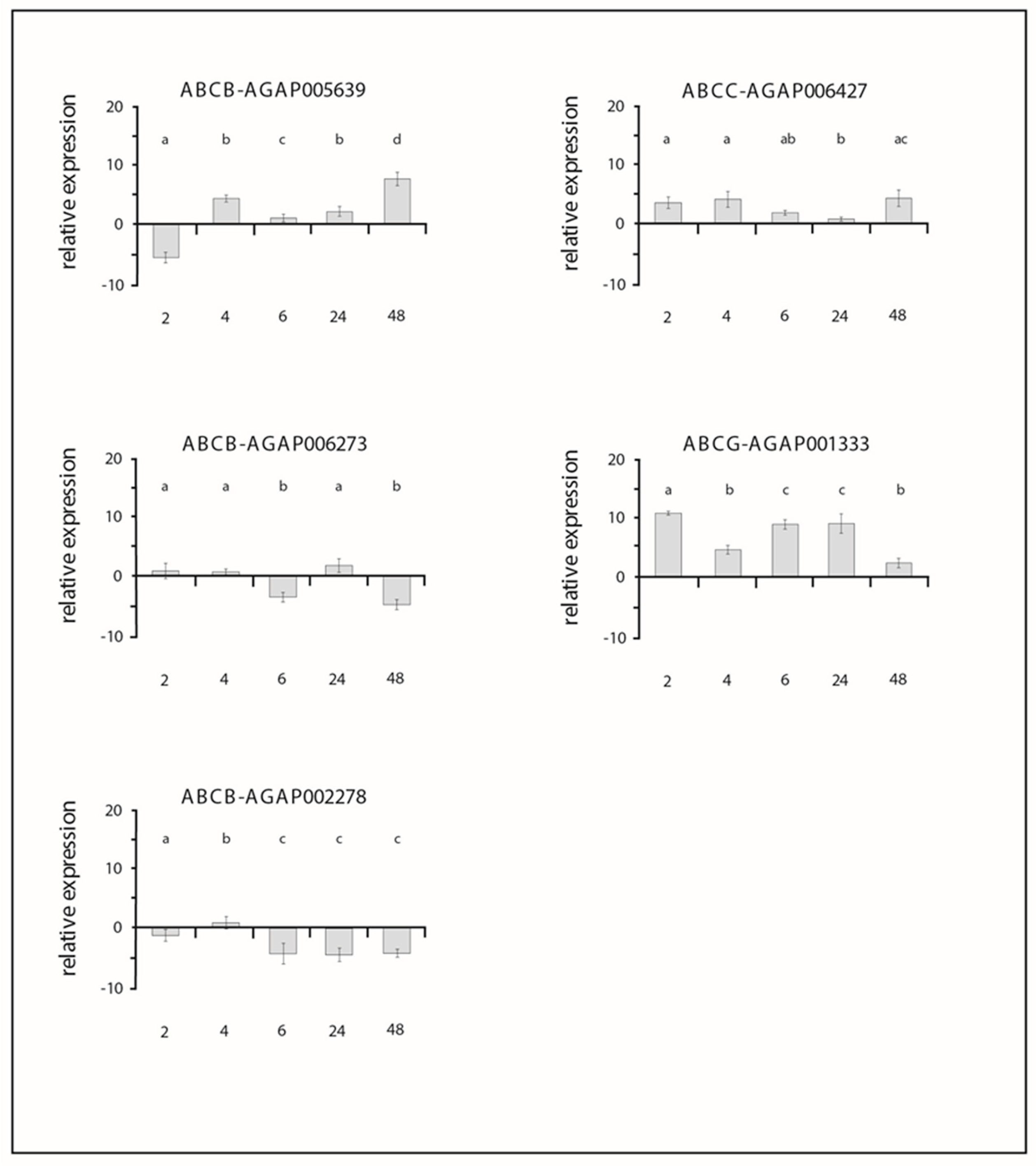
3. Results
3.1. Laboratory Bioassays
3.2. Expression Profiles of ABC Transporter Genes after Permethrin Exposure
4. Discussion
4.1. Involvement of ABC Transporters in Permethrin Detoxification
4.2. Dynamics of Gene Transcriptional Induction during Permethrin Exposure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Eliminating Malaria; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- McGraw, E.A.; O’Neill, S.L. Beyond insecticides: New thinking on an ancient problem. Nat. Rev. Microbiol. 2013, 11, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Wilke, A.B.; Marrelli, M.T. Paratransgenesis: A promising new strategy for mosquito vector control. Parasit. Vectors 2015, 8, 342. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Plan for Insecticide Resistance Management; World Health Organization: Geneva, Switzerland, 2012; p. 130. [Google Scholar]
- Liu, N. Insecticide resistance in mosquitoes: Impact, mechanisms and research directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef] [PubMed]
- Ranson, H.; Lissenden, N. Insecticide Resistance in African Anopheles Mosquitoes: A Worsening Situation that Needs Urgent Action to Maintain Malaria Control. Trends Parasitol. 2016, 32, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Chouaïbou, M.; Zivanovic, G.B.; Knox, T.B.; Jamet, H.P.; Bonfoh, B. Synergist bioassays: A simple method for initial metabolic resistance investigation of field Anopheles gambiae s.l. populations. Acta Trop. 2014, 130, 108–111. [Google Scholar] [CrossRef] [PubMed]
- Pohl, P.C.; Klafke, G.M.; Carvalho, D.D.; Martins, J.R.; Daffre, S.; da Silva Vaz, I. Jr.; Masuda, A. ABC transporter efflux pumps: A defence mechanism against ivermectin in Rhipicephalus (Boophilus) microplus. Int. J. Parasitol. 2011, 41, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.S.; Strycharz, J.P.; Baek, J.H.; Sun, W.; Kim, J.H.; Kang, J.S.; Pittendringh, B.R.; Lee, S.H.; Clarck, J.M. Brief exposures of human body lice to sublethal amounts of ivermectin over-transcribes detoxification genes involved in tolerance. Insect Mol. Biol. 2011, 20, 687–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueira-Mansur, J.; Ferreira-Pereira, A.; Mansur, J.F.; Franco, T.A.; Alvarenga, E.S.; Sorgine, M.H.; Neves, B.C.; Melo, A.C.; Leal, W.S.; Masuda, H.; et al. Silencing of P-glycoprotein increases mortality in temephos-treated Aedes aegypti larvae. Insect Mol. Biol. 2013, 226, 648–658. [Google Scholar] [CrossRef] [PubMed]
- Bona, A.C.; Chitolina, R.F.; Fermino, M.L.; de Castro Poncio, L.; Weiss, A.; Lima, J.B.; Paldi, N.; Bernardes, E.S.; Henen, J.; Maori, E. Larval application of sodium channel homologous dsRNA restores pyrethroid insecticide susceptibility in a resistant adult mosquito population. Parasit. Vectors 2016, 14, 397. [Google Scholar] [CrossRef] [PubMed]
- Buss, D.S.; Callaghan, A. Interaction of pesticides with p-glycoprotein and other ABC proteins: A survey of the possible importance to insecticide: Herbicide and fungicide resistance. Pestic. Biochem. Physiol. 2008, 90, 141–153. [Google Scholar] [CrossRef]
- Porretta, D.; Gargani, M.; Bellini, R.; Medici, A.; Punelli, F.; Urbanelli, S. Defence mechanism against insecticides temephos and diflubenzuron in the mosquito Aedes caspius: The P-glycoprotein efflux pumps. Med. Vet. Entomol. 2008, 22, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Dermauw, W.; van Leeuwen, T. The ABC gene family in arthropods: Comparative genomics and role in insecticide transport and resistance. Insect. Biochem. Mol. Biol. 2014, 45, 89–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porretta, D.; Epis, S.; Mastrantonio, V.; Ferrari, M.; Bellini, R.; Favia, G.; Urbanelli, S. How heterogeneous is the involvement of ABC transporters against insecticides? Acta Trop. 2016, 157, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Goldstone, J.; Hamdoun, A.M.; Cole, B.; Ashby, M.H.; Scally, M.; Dean, M.; Epel, D.; Hahn, M.E.; Stegeman, J.J. The chemical defensome: Environmental sensing and response genes in the Strongylocentrotus purpuratus genome. Dev. Biol. 2006, 300, 366–384. [Google Scholar] [CrossRef] [PubMed]
- Goldstone, J. Environmental Sensing and Response Genes in Cnidaria: The Chemical Defensome in the Sea Anemone Nematostella vectensis. Cell. Biol. Toxicol. 2008, 24, 483–502. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, C.E.W. Arms Race between Plants and Animals: Biotransformation System. In Stress Ecology: Environmental Stress as Ecological Driving Force and Key Player in Evolution; Steinberg, C.E.W., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 61–105. [Google Scholar] [CrossRef]
- De Marco, L.; Sassera, D.; Epis, S.; Mastrantonio, V.; Ferrari, M.; Ricci, I.; Comandatore, F.; Bandi, C.; Porretta, D.; Urbanelli, S. The choreography of the chemical defensome response to insecticide stress: Insights into the Anopheles stephensi transcriptome using RNA-Seq. Scien. Rep. 2017, 7, 41312. [Google Scholar] [CrossRef] [PubMed]
- Pignatelli, P.; Ingham, V.A.; Balabanidou, V.; Vontas, J.; Lycett, G.; Ranson, H. The Anopheles gambiae ATP-binding cassette transporter family: Phylogenetic analysis and tissue localization provide clues on function and role in insecticide resistance. Insect Mol. Biol. 2018, 27, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Pohl, P.C.; Klafke, G.M.; Júnior, J.R.; Martins, J.R.; da Silva Vaz, I., Jr.; Masuda, A. ABC transporters as a multidrug detoxification mechanism in Rhipicephalus (Boophilus) microplus. Paras. Res. 2012, 111, 2345–2351. [Google Scholar] [CrossRef] [PubMed]
- Cafarchia, C.; Porretta, D.; Mastrantonio, V.; Epis, S.; Sassera, D.; Iatta, R.; Immediato, D.; Ramos, R.A.; Lia, R.P.; Dantas-Torres, F.; et al. Potential role of ATP-binding cassette transporters against acaricides in the brown dog tick Rhipicephalus sanguineus sensu lato. Med. Vet. Entomol. 2015, 29, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Gujar, H.; Gordon, J.R.; Haynes, K.F.; Potter, M.F.; Palli, S.R. Bed bugs evolved unique adaptive strategy to resist pyrethroid insecticides. Sci. Rep. 2013, 3, 1456. [Google Scholar] [CrossRef] [PubMed]
- Buss, D.S.; McCaffery, A.R.; Callaghan, A. Evidence for p-glycoprotein modification of insecticide toxicity in mosquitoes of the Culex pipiens complex. Med. Vet. Entomol. 2002, 16, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Lima, E.P.; Goulart, M.O.F.; Neto, M.L.R. Evaluation of the role of ATP-binding cassette transporters as a defence mechanism against temephos in populations of Aedes aegypti. Mem. Inst. Oswaldo Cruz 2014, 109, 964–966. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Muñoz, V.; Nuñez-Acuña, G.; Gallardo-Escárate, C. Molecular characterization and transcription analysis of P-glycoprotein gene from the salmon louse Caligus rogercresseyi. J. Aquacult. Res. Dev. 2014, 5, 4. [Google Scholar] [CrossRef]
- Mastrantonio, V.; Ferrari, M.; Epis, S.; Negri, A.; Scuccimarra, G.; Montagna, M.; Favia, G.; Porretta, D.; Urbanelli, S.; Bandi, C. Gene expression modulation of ABC transporter genes in response to permethrin in adults of the mosquito malaria vector Anopheles stephensi. Acta Trop. 2017, 171, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Heckel, D.G. Learning the ABCs of Bt: ABC transporters and insect resistance to Bacillus thuringiensis provide clues to a crucial step in toxin mode of action. Pestic. Biochem. Physiol. 2012, 104, 103–110. [Google Scholar] [CrossRef]
- Bonizzoni, M.; Afrane, Y.; Dunn, W.A.; Atieli, F.K.; Zhou, G.; Zhong, D. Comparative transcriptome analyses of deltamethrin-resistant and -susceptible Anopheles gambiae mosquitoes from Kenya by RNA-Seq. PLoS ONE 2012, 7, e44607. [Google Scholar] [CrossRef] [PubMed]
- Yadouleton, A.; Martin, T.; Padonou, G.; Chandre, F.; Asidi, A.; Djogbenou, L.; Dabiré, R.; Aïkpon, R.; Boko, M.; Glitho, I.; et al. Cotton pest management practices and the selection of pyrethroid resistance in Anopheles gambiae population in Northern Benin. Parasit. Vectors 2011, 4, 60. [Google Scholar] [CrossRef] [PubMed]
- Bigoga, J.D.; Ndangoh, D.N.; Ambene, P.H.A.; Patchoké, S.; Fondjo, E.; Leke, R.G. Pyrethroid resistance in Anopheles gambiae from the rubber cultivated area of Niete, South Region of Cameroon. Acta Trop. 2012, 124, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Gnankiné, O.; Bassolé, I.H.N.; Chandre, F.; Glitho, I.; Akogbeto, M.; Dabiré, R.; Martin, T. Insecticide resistance in Bemisia tabaci Gennadius (Homoptera: Aleyrodidae) and Anopheles gambiae Giles (Diptera: Culicidae) could compromise the sustainability of malaria vector control strategies in West Africa. Acta Trop. 2013, 128, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Epis, S.; Porretta, D.; Mastrantonio, V.; Urbanelli, S.; Sassera, D.; Rossi, P. ABC transporters are involved in defense against permethrin insecticide in the malaria vector Anopheles stephensi. Parasit. Vectors 2014, 7, 349. [Google Scholar] [CrossRef] [PubMed]
- Epis, S.; Porretta, D.; Mastrantonio, V.; Comandatore, F.; Sassera, D.; de Marco, L. Temporal dynamics of the ABC transporter response to insecticide treatment: Insights from the malaria vector Anopheles stephensi. Sci. Rep. 2014, 4, 7435. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.M.; Toé, H.K.; Sanou, A.; Namountougou, M.; Hughes, A.; Diabaté, A.; Dabiré, R.; Simard, F.; Ranson, H. Additional selection for insecticide resistance in urban malaria vectors: DDT resistance in Anopheles arabiensis from Bobo-Dioulasso, Burkina Faso. PLoS ONE 2012, 7, e45995. [Google Scholar] [CrossRef] [PubMed]
- Merzendorfer, H. ABC Transporters and their role in protecting insects from pesticides and their metabolites. In Advances in Insect Physiology. Target Receptors in the Control of the Insect Pests: Part II; Cohen, E., Ed.; Academic Press: London, UK, 2014; pp. 1–73. [Google Scholar] [CrossRef]
- Tribble, N.D.; Burka, J.F.; Kibenge, F.S.B. Evidence for changes in the transcription levels of two putative P-glycoprotein genes in sea lice (Lepeophtheirus salmonis) in response to emamectin benzoate exposure. Mol. Biochem. Parasitol. 2007, 153, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Xie, W.; Yang, X.; Wang, S.; Wu, Q.; Li, R.; Pan, H.; Liu, B.; Shi, X.; Fang, Y.; et al. Transcriptomic and proteomic responses of sweetpotato whitefly, Bemisia tabaci, to thiamethoxam. PLoS ONE 2013, 8, e61820. [Google Scholar] [CrossRef] [PubMed]
- Fossog, T.B.; Kopya, E.; Ndo, C.; Menze-Djantio, B.; Costantini, C.; Njiokou, F.; Awono-Ambene, P.; Nkondjio, A.C. Water quality and Anopheles gambiae larval tolerance to pyrethroids in the cities of Douala and Yaoundé (Cameroon). J. Trop. Med. 2012, 429817. [Google Scholar] [CrossRef]
- Robertson, J.A.; Preisler, H.K.; Russell, R.M. Polo Plus: Probit and Logit Analysis; LeOra Software: Petaluma, CA, USA, 2003. [Google Scholar]
- Thornton, B.; Basu, C. Real-time PCR (qPCR) primer design using free online software. Biochem. Mol. Biol. Educ. 2011, 39, 145–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dönmez, Y.; Akhmetova, L.; İşeri, Ö.D.; Kars, M.D.; Gündüz, U. Effect of MDR modulators verapamil and promethazine on gene expression levels of MDR1 and MRP1 in doxorubicin-resistant MCF-7 cells. Cancer Chemother. Pharmacol. 2011, 67, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Jalali, A.; Ghasemian, S.; Najafzadeh, H.; Galehdari, H.; Seifi, M.R.; Zangene, F.; Dehdardargahi, S. Verapamil and rifampin effect on p-glycoprotein expression in hepatocellular carcinoma. J. Nat. Pharm. Prod. 2014, 10, e17741. [Google Scholar] [CrossRef]
- Sulová, Z.; Macejová, D.; Seres, M.; Sedlák, J.; Brtko, J.; Breier, A. Combined treatment of P-gp-positive L1210/VCR cells by verapamil and all-trans retinoic acid induces down-regulation of P-glycoprotein expression and transport activity. Toxicol. In Vitro 2008, 22, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Li, T.; Reid, W.R.; Yang, T.; Zhang, L. Multiple cytochrome P450 genes: Their constitutive overexpression and permethrin induction in insecticide resistant mosquitoes, Culex quinquefasciatus. PLoS ONE 2011, 6, e23403. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Li, T.; Zhang, L.; Gao, X.; Liu, N. Permethrin induction of multiple cytochrome P450 genes in insecticide-resistant mosquitoes, Culex quinquefasciatus. Int. J. Biol. Sci. 2013, 9, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Doganlar, O.; Doganlar, Z.B. Responses of antioxidant enzymes and heat shock proteins in Drosophila to treatment with a pesticide mixture. Arch. Biol. Sci. Belgrade 2015, 67, 869–876. [Google Scholar] [CrossRef]
- Fang, F.; Wang, W.; Zhang, D.; Lv, Y.; Zhou, D.; Ma, L.; Shen, B.; Sun, Y.; Zhu, C. The cuticle proteins: A putative role for deltamethrin resistance in Culex pipiens pallens. Parasitol. Res. 2015, 114, 4421–4429. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Vector Base Sequence ID | ABC Sub-Family | Forward 3’-5’ | Reverse 3’-5’ | PCR Product Size (bp) |
|---|---|---|---|---|
| AGAP005639 | ABCB | TTCATCACGAAACTACCGAAC | GTCCCTTACTTGCTCGCT | 204 |
| AGAP006273 | ABCB | CACGCTGGGCTATCAGTA | AAAACTTCCACCAATCGAAACG | 118 |
| AGAP002278 | ABCB | AAAGGTGACAGAGAGGTGTAGGAAA | ACGCCATGCACTAAACTATCACATT | 104 |
| AGAP006427 | ABCC | AAAGTGTTCTACGGCATGGTGAAG | CAGCCTCCTTAATCGGTTTCAGTTT | 108 |
| AGAP001333 | ABCG | GTCTCCTGTCGTTGTAGTTTT | CGTAACAGAAACATCGTCCATT | 174 |
| AGAP010592 | rps7 | GGCGATCATCATCTACGTGC | GTAGCTGCTGCAAACTTCGG | 459 |
| AGAP000651 | act 5C | TCTGGCACCACACGTTCTAC | CAGGTAGTCGGTGAGATCGC | 313 |
| Insecticide | Slope (±SE) | LD50 (95% CI) | SR (95% CI) | χ2 (df) | ||
|---|---|---|---|---|---|---|
| Goodness-of-Fit | Equality | Parallelism | ||||
| permethrin | 1.514 (0.131) | 297.84 ppb (248.5–368.18) | 2.467 (4) | 317 (2) * | 11.40 (1) * | |
| permethrin + verapamil | 2.259 (0.181) | 18.69 ppb (15.89–21.81) | 15.94 (12.37–20.53) | 13.37 (5) | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mastrantonio, V.; Ferrari, M.; Negri, A.; Sturmo, T.; Favia, G.; Porretta, D.; Epis, S.; Urbanelli, S. Insecticide Exposure Triggers a Modulated Expression of ABC Transporter Genes in Larvae of Anopheles gambiae s.s. Insects 2019, 10, 66. https://doi.org/10.3390/insects10030066
Mastrantonio V, Ferrari M, Negri A, Sturmo T, Favia G, Porretta D, Epis S, Urbanelli S. Insecticide Exposure Triggers a Modulated Expression of ABC Transporter Genes in Larvae of Anopheles gambiae s.s. Insects. 2019; 10(3):66. https://doi.org/10.3390/insects10030066
Chicago/Turabian StyleMastrantonio, Valentina, Marco Ferrari, Agata Negri, Tommaso Sturmo, Guido Favia, Daniele Porretta, Sara Epis, and Sandra Urbanelli. 2019. "Insecticide Exposure Triggers a Modulated Expression of ABC Transporter Genes in Larvae of Anopheles gambiae s.s." Insects 10, no. 3: 66. https://doi.org/10.3390/insects10030066