
Seaweed Supplements Normalise Metabolic, Cardiovascular and Liver Responses in High-Carbohydrate, High-Fat Fed Rats

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Nutritional Composition of Seaweeds
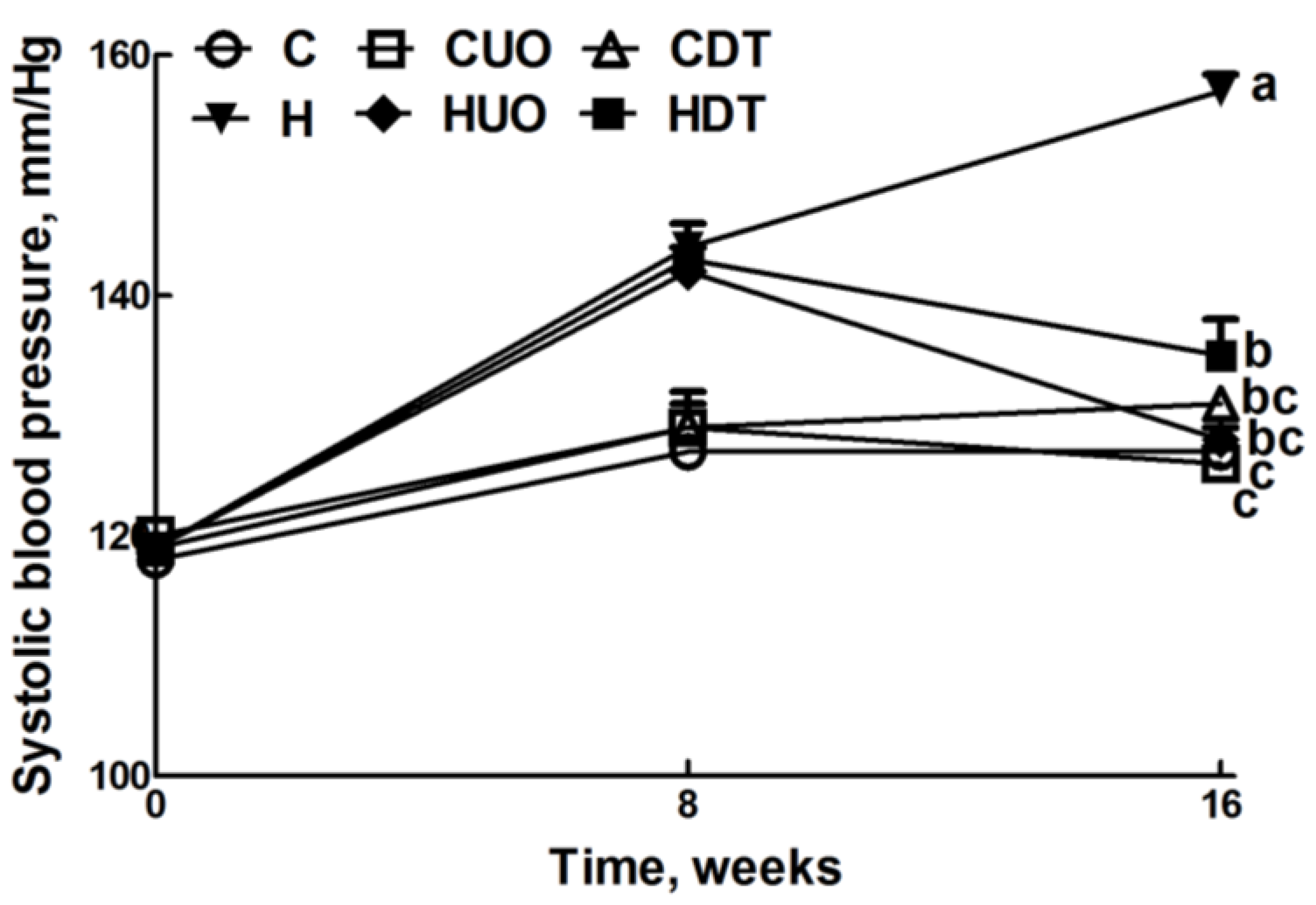
2.2. Metabolic Variables

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | C | CUO | CDT | H | HUO | HDT | Diet | Treatment | Interaction |
|---|---|---|---|---|---|---|---|---|---|
| Food intake, g/day | 35.7 ± 3.7 a | 33.6 ± 3.6 a | 32.2 ± 2.4 a | 22.0 ± 2.0 b | 21.6 ± 0.8 b | 21.9 ± 1.3 b | <0.0001 | 0.78 | 0.81 |
| Water intake, mL/day | 32.0 ± 3.4 a | 33.9 ± 2.8 a | 33.6 ± 2.4 a | 20.0 ± 1.8 b | 24.8 ± 0.8 b | 20.9 ± 1.7 b | <0.0001 | 0.37 | 0.73 |
| Energy intake, kJ/day | 396.7 ± 20.2 b | 398.9 ± 11.1 b | 406.6 ± 11.3 b | 469.9 ± 9.3 a | 483.2 ± 15.2 a | 478.2 ± 4.1 a | <0.0001 | 0.75 | 0.86 |
| Feed conversion efficiency, % | 2.8 ± 0.3 ab | 2.5 ± 0.3 b | 4.1 ± 0.4 a | 2.9 ± 0.3 ab | 2.9 ± 0.2 ab | 3.6 ± 0.5 ab | 1.00 | 0.004 | 0.44 |
| Body weight gain, % | 11.1 ± 1.1 b | 9.9 ± 1.1 b | 16.7 ± 1.5 a | 12.8 ± 1.4 ab | 13.8 ± 1.2 ab | 17.3 ± 2.2 a | 0.10 | 0.001 | 0.54 |
| Variable | C | CUO | CDT | H | HUO | HDT | P-Value | ||
|---|---|---|---|---|---|---|---|---|---|
| Diet | Treatment | Interaction | |||||||
| Bone mineral content, g | 12.4 ± 0.3 c | 12.4 ± 0.5 c | 13.5 ± 0.4 bc | 15.1 ± 0.5 a | 14.3 ± 0.4 ab | 15.7 ± 0.4 a | <0.0001 | 0.017 | 0.64 |
| Bone mineral density, g/cm2 | 0.162 ± 0.003 | 0.163 ± 0.003 | 0.164 ± 0.003 | 0.167 ± 0.002 | 0.161 ± 0.002 | 0.162 ± 0.002 | 0.88 | 0.63 | 0.30 |
| Total lean mass, g | 319.1 ± 10.3 ab | 302.4 ± 2.5 bc | 333.2 ± 6.6 a | 271.4 ± 8.4 c | 292.3 ± 11.8 bc | 284.4 ± 3.9 c | <0.0001 | 0.23 | 0.04 |
| Total body fat mass, g | 85.5 ± 7.4 c | 103.8 ± 9.8 c | 100.1 ± 7.9 c | 201.6 ± 10.9 a | 153.1 ± 14.6 b | 190.0 ± 18.9 a | <0.0001 | 0.32 | 0.026 |
| Abdominal circumference, cm | 19.6 ± 0.4 b | 18.8 ± 0.2 b | 19.6 ± 0.2 b | 22.0 ± 0.4 a | 21.4 ± 0.2 a | 22.2 ± 0.6 a | <0.0001 | 0.08 | 0.95 |
| Glucose AUC, mmol/L/120 min | 637.6 ± 8.4 c | 674.9 ± 17.6 bc | 700.7 ± 22.0 b | 809.7 ± 12.9 a | 703.5 ± 18.8 b | 728.6 ± 12.2 b | <0.0001 | 0.09 | <0.0001 |
| Insulin AUC, mmol/L/120 min | 169.5 ± 22.8 c | 226.8 ± 6.2 bc | 228.7 ± 13.9 bc | 391.8 ± 16.2 a | 226.5 ± 18.3 bc | 254.5 ± 33.9 b | <0.0001 | 0.03 | <0.0001 |
| Retroperitoneal fat, mg/mm tibial length | 138.8 ± 11.1 b | 127.8 ± 10.6 b | 150.4 ± 9.3 b | 331.5 ± 20.9 a | 303.3 ± 16.9 a | 322.0 ± 18.8 a | <0.0001 | 0.32 | 0.76 |
| Epididymal fat, mg/mm tibial length | 105.3 ± 5.7 b | 91.3 ± 4.8 b | 115.3 ± 8.4 b | 170.4 ± 11.4 a | 200.3 ± 15.4 a | 197.1 ± 16.7 a | <0.0001 | 0.26 | 0.15 |
| Omental fat, mg/mm tibial length | 59.7 ± 5.7 b | 63.1 ± 4.6 b | 74.3 ± 6.6 b | 131.6 ± 10.6 a | 125.4 ± 12.3 a | 115.3 ± 14.4 a | <0.0001 | 0.99 | 0.27 |
| Total abdominal fat, mg/mm tibial length | 292.7 ± 19.6 b | 268.9 ± 26.0 b | 340.0 ± 23.1 b | 670.1 ± 46.9 a | 628.9 ± 42.4 a | 634.4 ± 40.2 a | <0.0001 | 0.49 | 0.45 |
| Visceral adiposity index, % | 3.7 ± 0.3 b | 3.0 ± 0.3 b | 3.9 ± 0.3 b | 6.9 ± 0.5 a | 7.0 ± 0.4 a | 7.3 ± 0.7 a | <0.0001 | 0.41 | 0.65 |
| Plasma NEFA, mmol/L | 1.47 ± 0.18 c | 1.46 ± 0.28 c | 1.55 ± 0.09 c | 2.78 ± 0.28 b | 3.73 ± 0.29 a | 2.72 ± 0.47 b | <0.0001 | 0.18 | 0.12 |
| Plasma triglycerides, mmol/L | 0.43 ± 0.07 c | 0.41 ± 0.07 c | 0.54 ± 0.07 c | 1.29 ± 0.19 a | 1.17 ± 0.14 ab | 0.80 ± 0.19 bc | <0.0001 | 0.38 | 0.07 |
| Plasma total cholesterol, mmol/L | 1.52 ± 0.09 b | 1.51 ± 0.04 b | 1.53 ± 0.05 b | 1.98 ± 0.05 a | 1.87 ± 0.19 ab | 1.65 ± 0.09 b | 0.0002 | 0.26 | 0.20 |
| Variable | C | CUO | CDT | H | HUO | HDT | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|
| Diet | Treatment | Interaction | |||||||
| LV + septum, mg/mm tibial length | 17.1 ± 0.3 b | 18.9 ± 0.4 ab | 20.5 ± 1.0 a | 17.2 ± 0.5 b | 17.3 ± 0.3 b | 18.1 ± 1.0 b | 0.023 | 0.010 | 0.18 |
| RV wet weight, mg/mm tibial length | 2.2 ± 0.2 ab | 2.7 ± 0.2 ab | 2.9 ± 0.4 a | 2.2 ± 0.1 ab | 2.0 ± 0.1 ab | 1.9 ± 0.2 b | 0.004 | 0.67 | 0.10 |
| Heart wet weight, mg/mm tibial length | 19.3 ± 0.4 b | 20.5 ± 0.5 b | 23.3 ± 1.4 a | 19.4 ± 0.5 b | 19.2 ± 0.5 b | 19.3 ± 0.5 b | 0.006 | 0.030 | 0.025 |
| Systolic blood pressure, mmHg | 127 ± 2 c | 126 ± 3 c | 131 ± 1 bc | 157 ± 1 a | 128 ± 1bc | 135 ± 3 b | <0.0001 | <0.0001 | <0.0001 |
| SBP:LVIDs | 35.2 ± 1.8 b | 29.4 ± 2.0 c | 30.4 ± 1.9 c | 47.3 ± 3.9 a | 29.2 ± 0.9 b | 40.1 ± 1.6 b | 0.0006 | <0.0001 | 0.033 |
| SBP:systolic volume | 3096 ± 525 ab | 1731 ± 363 b | 1716 ± 340 b | 4818 ± 1129 a | 1497 ± 137 b | 3690 ± 458 ab | 0.035 | 0.003 | 0.18 |
| ESS:LVIDs | 2.02 ± 0.07 b | 2.10 ± 0.06 b | 2.09 ± 0.08 b | 2.44 ± 0.09 a | 2.17 ± 0.07 b | 2.02 ± 0.08 b | 0.032 | 0.09 | 0.008 |
| Diastolic stiffness κ | 23.9 ± 1.7 b | 23.3 ± 0.4 b | 23.0 ± 0.6 b | 29.8 ± 2.2 a | 25.1 ± 0.9 b | 23.4 ± 1.0 b | 0.016 | 0.023 | 0.11 |
| Variable | C | CUO | CDT | H | HUO | HDT | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|
| Diet | Treatment | Interaction | |||||||
| Liver weight | 215.7 ± 6.8 b | 236.6 ± 13.9 b | 229.9 ± 7.9 b | 277.5 ± 13.6 a | 301.2 ± 10.2 a | 269.3 ± 18.0 a | <0.0001 | 0.17 | 0.55 |
| Plasma ALT activity, U/L | 28.2 ± 2.4 b | 32.4 ± 1.8 b | 36.5 ± 3.1 b | 47.4 ± 4.1 a | 30.2 ± 2.6 b | 31.9 ± 2.9 b | 0.26 | 0.10 | 0.0002 |
| Plasma AST activity, U/L | 71.7 ± 5.3 b | 84.1 ± 2.6 b | 85.5 ± 3.3 b | 102.4 ± 3.4 a | 70.8 ± 4.5 b | 89.5 ± 3.5 b | 0.10 | 0.05 | <0.0001 |
| Plasma Na+, mmol/L | 143 ± 1 | 143 ± 1 | 141 ± 1 | 143 ± 0 | 140 ± 1 | 140 ± 1 | 0.07 | 0.03 | 0.24 |
| Plasma K+, mmol/L | 5.55 ± 0.38 | 5.94 ± 0.33 | 6.43 ± 0.31 | 6.03 ± 0.64 | 6.75 ± 1.04 | 5.20 ± 0.48 | 0.97 | 0.57 | 0.19 |
| Plasma Mg2+, mmol/L | 0.78 ± 0.03 | 0.86 ± 0.04 | 0.83 ± 0.02 | 0.79 ± 0.03 | 0.89 ± 0.04 | 0.81 ± 0.03 | 0.80 | 0.027 | 0.74 |

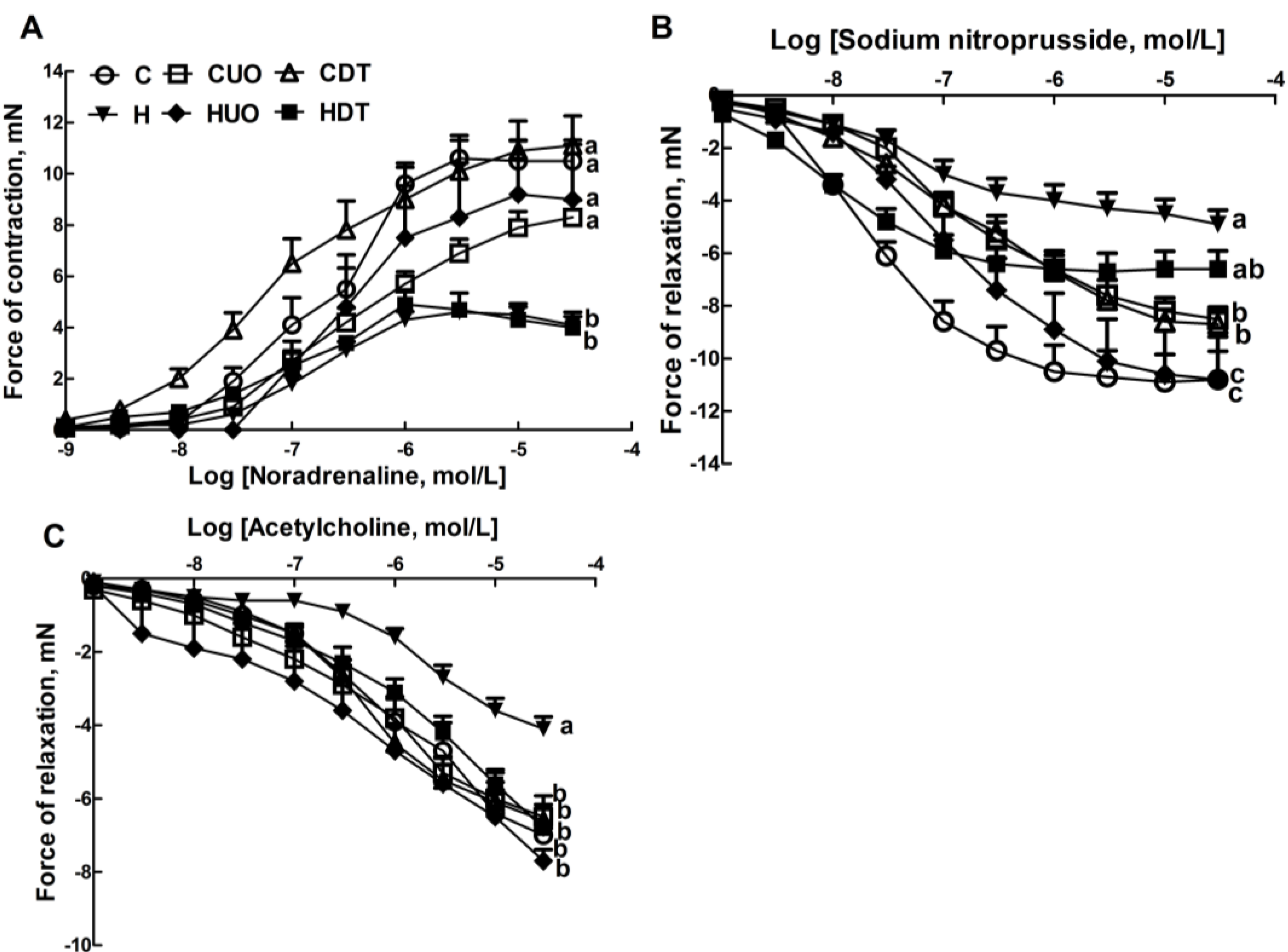
2.3. Cardiovascular Structure and Function


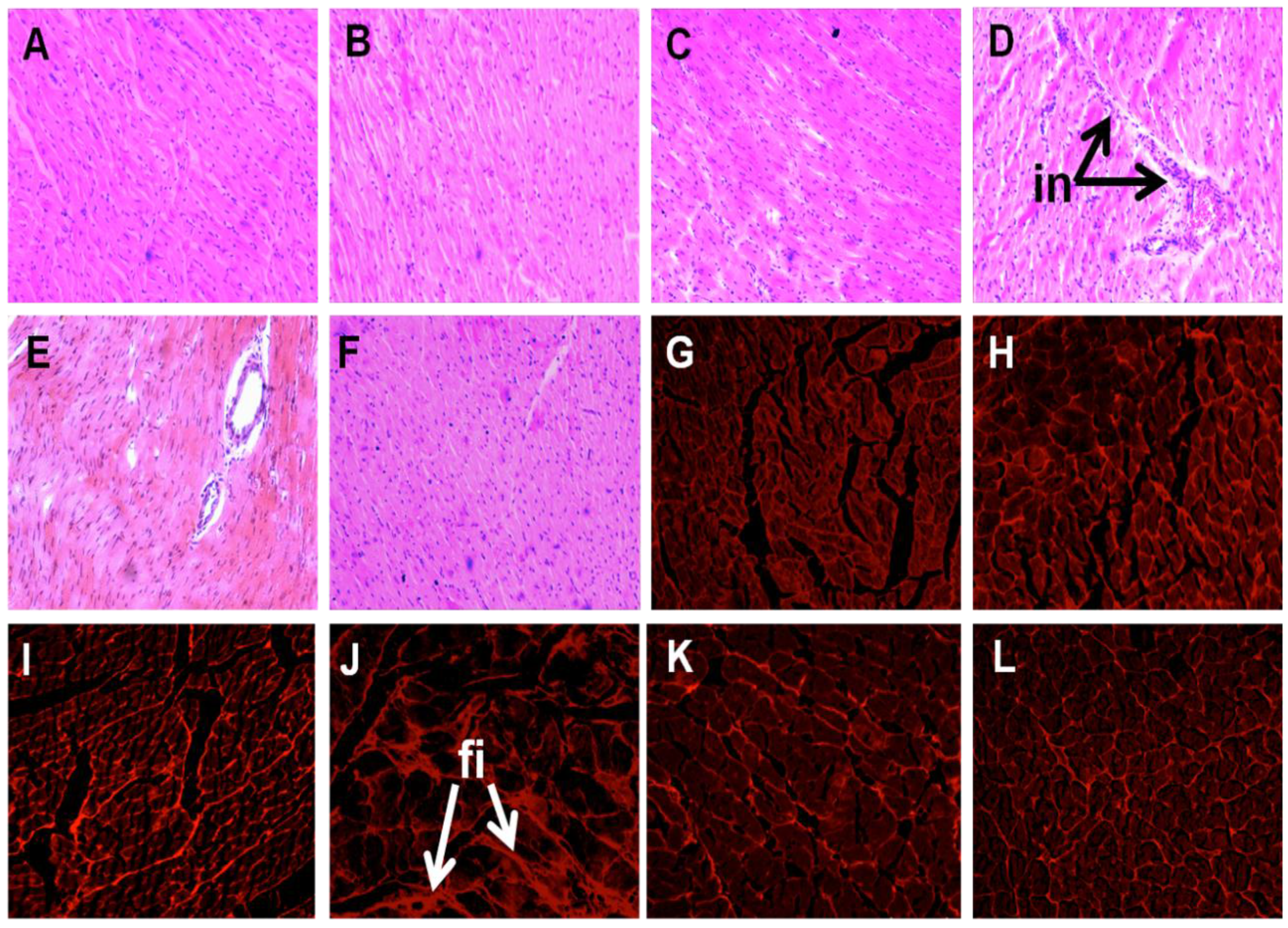
2.4. Liver Structure and Function

3. Discussion
4. Experimental Section
4.1. Resources: Algae, Diet Components, Rats

4.2. Cardiovascular Measurements
4.3. Organ Weights
4.4. Histology
4.5. Plasma Biochemistry
4.6. Statistical Analysis
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H. Obesity and cardiovascular disease: Pathophysiology, evaluation, and effect of weight loss. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 968–976. [Google Scholar] [CrossRef] [PubMed]
- Lavie, C.J.; Milani, R.V.; Ventura, H.O. Obesity and cardiovascular disease: Risk factor, paradox, and impact of weight loss. J. Am. Coll. Cardiol. 2009, 53, 1925–1932. [Google Scholar] [CrossRef] [PubMed]
- Munsters, M.J.; Saris, W.H. Body weight regulation and obesity: Dietary strategies to improve the metabolic profile. Annu. Rev. Food Sci. Technol. 2014, 5, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Chatzigeorgiou, A.; Kandaraki, E.; Papavassiliou, A.G.; Koutsilieris, M. Peripheral targets in obesity treatment: A comprehensive update. Obes. Rev. 2014, 15, 487–503. [Google Scholar] [CrossRef] [PubMed]
- Gadde, K.M. Current pharmacotherapy for obesity: Extrapolation of clinical trials data to practice. Expert Opin. Pharmacother. 2014, 15, 809–822. [Google Scholar] [CrossRef] [PubMed]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [PubMed]
- MacArtain, P.; Gill, C.I.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional value of edible seaweeds. Nutr. Rev. 2007, 65, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.; Hashim, S.N.; Rahman, H.A. Seaweeds: A sustainable functional food for complementary and alternative therapy. Trends Food Sci. Tech. 2012, 23, 83–96. [Google Scholar] [CrossRef]
- Mayakrishnan, V.; Kannappan, P.; Abdullah, N.; Ahmed, A.B.A. Cardioprotective activity of polysaccharides derived from marine algae: An overview. Trends Food Sci. Technol. 2013, 30, 98–104. [Google Scholar] [CrossRef]
- Lahaye, M. Marine algae as sources of fibres: Determination of soluble and insoluble dietary fibre contents in some ‘sea vegetables’. J. Sci. Food Agric. 1991, 54, 587–594. [Google Scholar] [CrossRef]
- Dawczynski, C.; Schubert, R.; Jahreis, G. Amino acids, fatty acids, and dietary fibre in edible seaweed products. Food Chem. 2007, 103, 891–899. [Google Scholar] [CrossRef]
- Jakobsdottir, G.; Nyman, M.; Fak, F. Designing future prebiotic fiber to target metabolic syndrome. Nutrition 2014, 30, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Gosch, B.J.; Magnusson, M.; Paul, N.A.; de Nys, R. Total lipid and fatty acid composition of seaweeds for the selection of species for oil-based biofuel and bioproducts. GCB Bioenergy 2012, 4, 919–930. [Google Scholar] [CrossRef]
- Pengzhan, Y.; Ning, L.; Xiguang, L.; Gefei, Z.; Quanbin, Z.; Pengcheng, L. Antihyperlipidemic effects of different molecular weight sulfated polysaccharides from Ulva pertusa (Chlorophyta). Pharmacol. Res. 2003, 48, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Paul, V.J.; Luesch, H. Seaweed extracts and unsaturated fatty acid constituents from the green alga Ulva lactuca as activators of the cytoprotective Nrf2-ARE pathway. Biol. Med. 2013, 57, 141–153. [Google Scholar]
- Ramirez-Higuera, A.; Quevedo-Corona, L.; Paniagua-Castro, N.; Chamorro-Ceballos, G.; Milliar-Garcia, A.; Jaramillo-Flores, M. Antioxidant enzymes gene expression and antihypertensive effects of seaweeds Ulva linza and Lessonia trabeculata in rats fed a high-fat and high-sucrose diet. J. Appl. Phycol. 2014, 26, 597–605. [Google Scholar] [CrossRef]
- Lawton, R.J.; Mata, L.; de Nys, R.; Paul, N.A. Algal bioremediation of waste waters from land-based aquaculture using Ulva: Selecting target species and strains. PLoS One 2013, 8, e77344. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, M.; Mata, L.; de Nys, R.; Paul, N.A. Biomass, lipid and fatty acid production in large-scale cultures of the marine macroalga Derbesia tenuissima (Chlorophyta). Mar. Biotechnol. (NY) 2014, 16, 456–464. [Google Scholar]
- Lee, H.J.; Kim, Y.A.; Lee, J.I.; Lee, B.J.; Seo, Y. Screening of Korean marine plants extracts for inhibitory activity on protein tyrosine phosphatase 1B. J. Appl. Biol. Chem. 2007, 57, 74–77. [Google Scholar]
- Panchal, S.K.; Poudyal, H.; Iyer, A.; Nazer, R.; Alam, M.A.; Diwan, V.; Kauter, K.; Sernia, C.; Campbell, F.; Ward, L.; et al. High-carbohydrate, high-fat diet-induced metabolic syndrome and cardiovascular remodeling in rats. J. Cardiovasc. Pharmacol. 2011, 57, 611–624. [Google Scholar]
- Panchal, S.K.; Poudyal, H.; Waanders, J.; Brown, L. Coffee extract attenuates changes in cardiovascular and hepatic structure and function without decreasing obesity in high-carbohydrate, high-fat diet-fed male rats. J. Nutr. 2012, 142, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Poudyal, H.; Panchal, S.K.; Ward, L.C.; Brown, L. Effects of ALA, EPA and DHA in high-carbohydrate, high-fat diet-induced metabolic syndrome in rats. J. Nutr. Biochem. 2013, 24, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Satija, A.; Hu, F. Cardiovascular benefits of dietary fiber. Curr. Atheroscler. Rep. 2012, 14, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Slavin, J.L. Position of the American Dietetic Association: Health implications of dietary fiber. J. Am. Diet. Ass. 2008, 108, 1716–1731. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Bocanegra, A.; Bastida, S.; Benedi, J.; Rodenas, S.; Sanchez-Muniz, F.J. Characteristics and nutritional and cardiovascular-health properties of seaweeds. J. Med. Food 2009, 12, 236–258. [Google Scholar] [CrossRef] [PubMed]
- Jensen, G.M.; Pedersen, C.; Kristensen, M.; Frost, G.; Astrup, A. Review: Efficacy of alginate supplementation in relation to appetite regulation and metabolic risk factors: Evidence from animal and human studies. Obes. Rev. 2013, 14, 129–144. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, M.; Knudsen, K.E.; Jorgensen, H.; Oomah, D.; Bugel, S.; Toubro, S.; Tetens, I.; Astrup, A. Linseed dietary fibers reduce apparent digestibility of energy and fat and weight gain in growing rats. Nutrients 2013, 5, 3287–3298. [Google Scholar] [CrossRef] [PubMed]
- Domozych, D.S.; Ciancia, M.; Fangel, J.U.; Mikkelsen, M.D.; Ulvskov, P.; Willats, W.G. The cell walls of green algae: A journey through evolution and diversity. Front. Plant Sci. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: Metabolic and health benefits. Br. J. Nutr. 2010, 104 (Suppl. 2), S1–S63. [Google Scholar]
- Backhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [PubMed]
- Nagai, N.; Ito, Y. Effect of magnesium ion supplementation on obesity and diabetes mellitus in Otsuka Long-Evans Tokushima Fatty (OLETF) rats under excessive food intake. J. Oleo Sci. 2013, 62, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Volpe, S.L. Magnesium, the metabolic syndrome, insulin resistance, and type 2 diabetes mellitus. Crit. Rev. Food Sci. Nutr. 2008, 48, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Sales, C.H.; Santos, A.R.; Cintra, D.E.; Colli, C. Magnesium-deficient high-fat diet: Effects on adiposity, lipid profile and insulin sensitivity in growing rats. Clin. Nutr. 2013, 33, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Zicha, J.; Dobesova, Z.; Behuliak, M.; Kunes, J.; Vaneckova, I. Preventive dietary potassium supplementation in young salt-sensitive Dahl rats attenuates development of salt hypertension by decreasing sympathetic vasoconstriction. Acta Physiol. (Oxf.) 2011, 202, 29–38. [Google Scholar] [CrossRef]
- Nobre, J.L.; Lisboa, P.C.; Santos-Silva, A.P.; Lima, N.S.; Manhaes, A.C.; Nogueira-Neto, J.F.; Cabanelas, A.; Pazos-Moura, C.C.; Moura, E.G.; de Oliveira, E. Calcium supplementation reverts central adiposity, leptin, and insulin resistance in adult offspring programed by neonatal nicotine exposure. J. Endocrinol. 2011, 210, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.; Wang, Y.; Sun, J.; Sun, W.; Tan, Y.; Cai, L.; Zheng, Y.; Su, G.; Liu, Q. Zinc protects against diabetes-induced pathogenic changes in the aorta: Roles of metallothionein and nuclear factor (erythroid-derived 2)-like 2. Cardiovasc. Diabetol. 2013, 12. [Google Scholar] [CrossRef]
- Whelton, S.P.; Hyre, A.D.; Pedersen, B.; Yi, Y.; Whelton, P.K.; He, J. Effect of dietary fiber intake on blood pressure: A meta-analysis of randomized, controlled clinical trials. J. Hypertens. 2005, 23, 475–481. [Google Scholar] [CrossRef] [PubMed]
- King, D.E.; Mainous, A.G.; Egan, B.M.; Woolson, R.F.; Geesey, M.E. Fiber and C-reactive protein in diabetes, hypertension, and obesity. Diabetes Care 2005, 28, 1487–1489. [Google Scholar] [PubMed]
- Gregor, M.F.; Hotamisligil, G.S. Inflammatory mechanisms in obesity. Annu. Rev. Immunol. 2011, 29, 415–445. [Google Scholar] [CrossRef] [PubMed]
- Ajani, U.A.; Ford, E.S.; Mokdad, A.H. Dietary fiber and C-reactive protein: Findings from National Health and Nutrition Examination Survey data. J. Nutr. 2004, 134, 1181–1185. [Google Scholar] [PubMed]
- Kumar, S.A.; Brown, L. Seaweeds as potential therapeutic interventions for the metabolic syndrome. Rev. Endocr. Metab. Disord. 2013, 14, 299–308. [Google Scholar] [CrossRef]
- Galvez, J.; Rodríguez-Cabezas, M.E.; Zarzuelo, A. Effects of dietary fiber on inflammatory bowel disease. Mol. Nutr. Food Res. 2005, 49, 601–608. [Google Scholar] [CrossRef] [PubMed]
- De Simone, G.; di Lorenzo, L.; Costantino, G.; Buonissimo, S.; Moccia, D. Echocardiographic indexes of left ventricular contractility. Effect of load manipulation in arterial hypertension. Jpn. Heart J. 1988, 29, 151–160. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, S.A.; Magnusson, M.; Ward, L.C.; Paul, N.A.; Brown, L. Seaweed Supplements Normalise Metabolic, Cardiovascular and Liver Responses in High-Carbohydrate, High-Fat Fed Rats. Mar. Drugs 2015, 13, 788-805. https://doi.org/10.3390/md13020788
Kumar SA, Magnusson M, Ward LC, Paul NA, Brown L. Seaweed Supplements Normalise Metabolic, Cardiovascular and Liver Responses in High-Carbohydrate, High-Fat Fed Rats. Marine Drugs. 2015; 13(2):788-805. https://doi.org/10.3390/md13020788
Chicago/Turabian StyleKumar, Senthil Arun, Marie Magnusson, Leigh C. Ward, Nicholas A. Paul, and Lindsay Brown. 2015. "Seaweed Supplements Normalise Metabolic, Cardiovascular and Liver Responses in High-Carbohydrate, High-Fat Fed Rats" Marine Drugs 13, no. 2: 788-805. https://doi.org/10.3390/md13020788