trans-Caryophyllene, a Natural Sesquiterpene, Causes Tracheal Smooth Muscle Relaxation through Blockade of Voltage-Dependent Ca2+ Channels


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
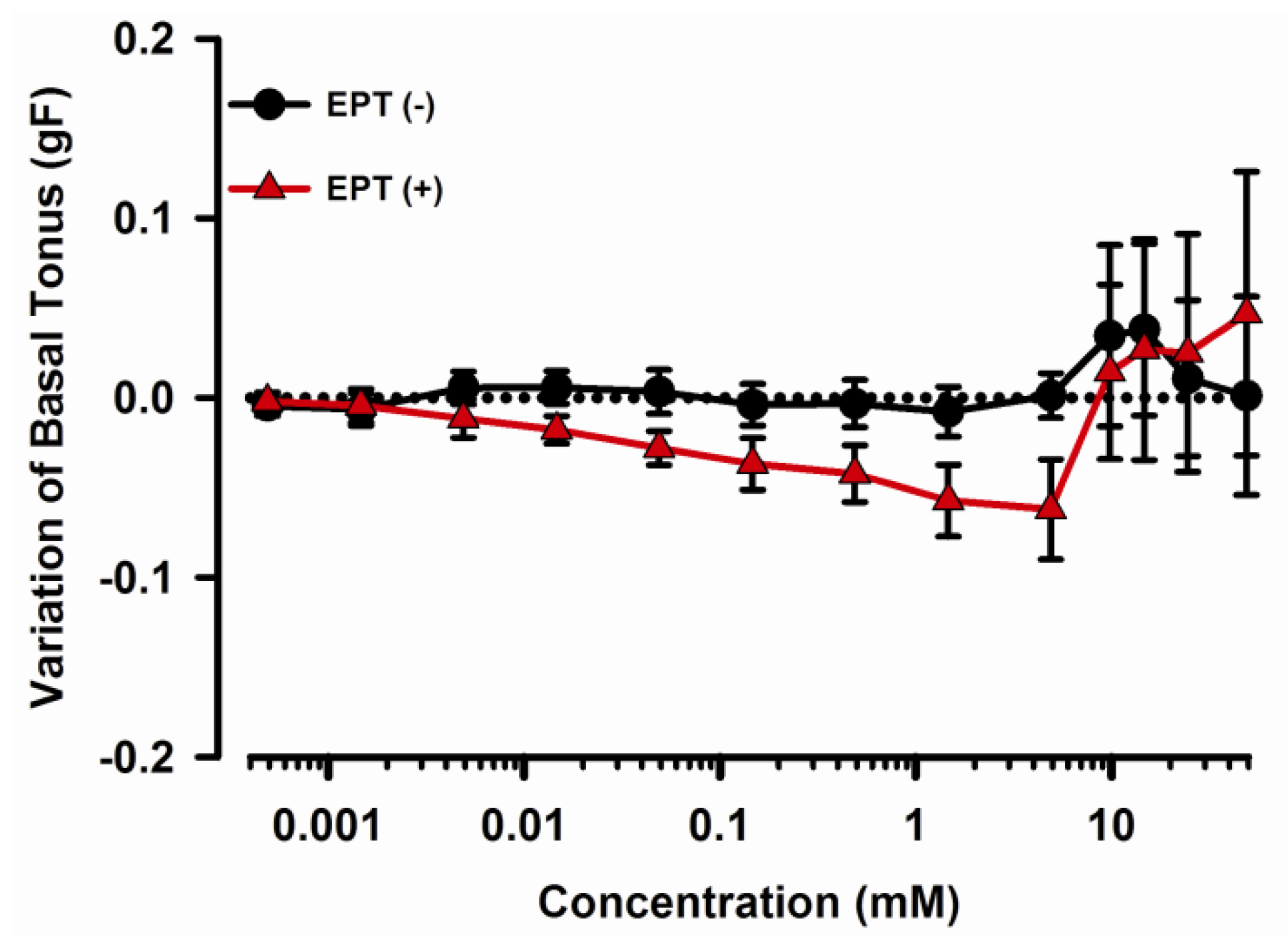
2.1. Effects of trans-Caryophyllene on Basal Tone of Isolated Rat Trachea with or without Epithelium
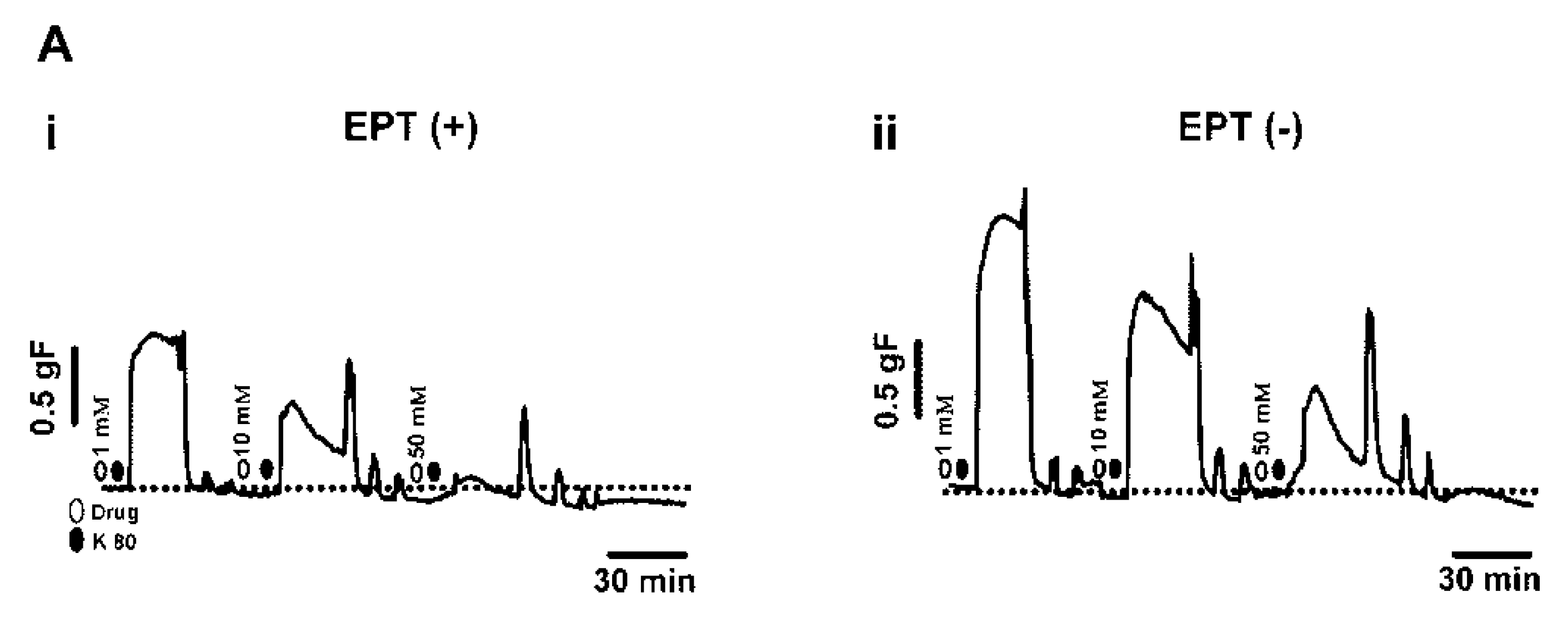
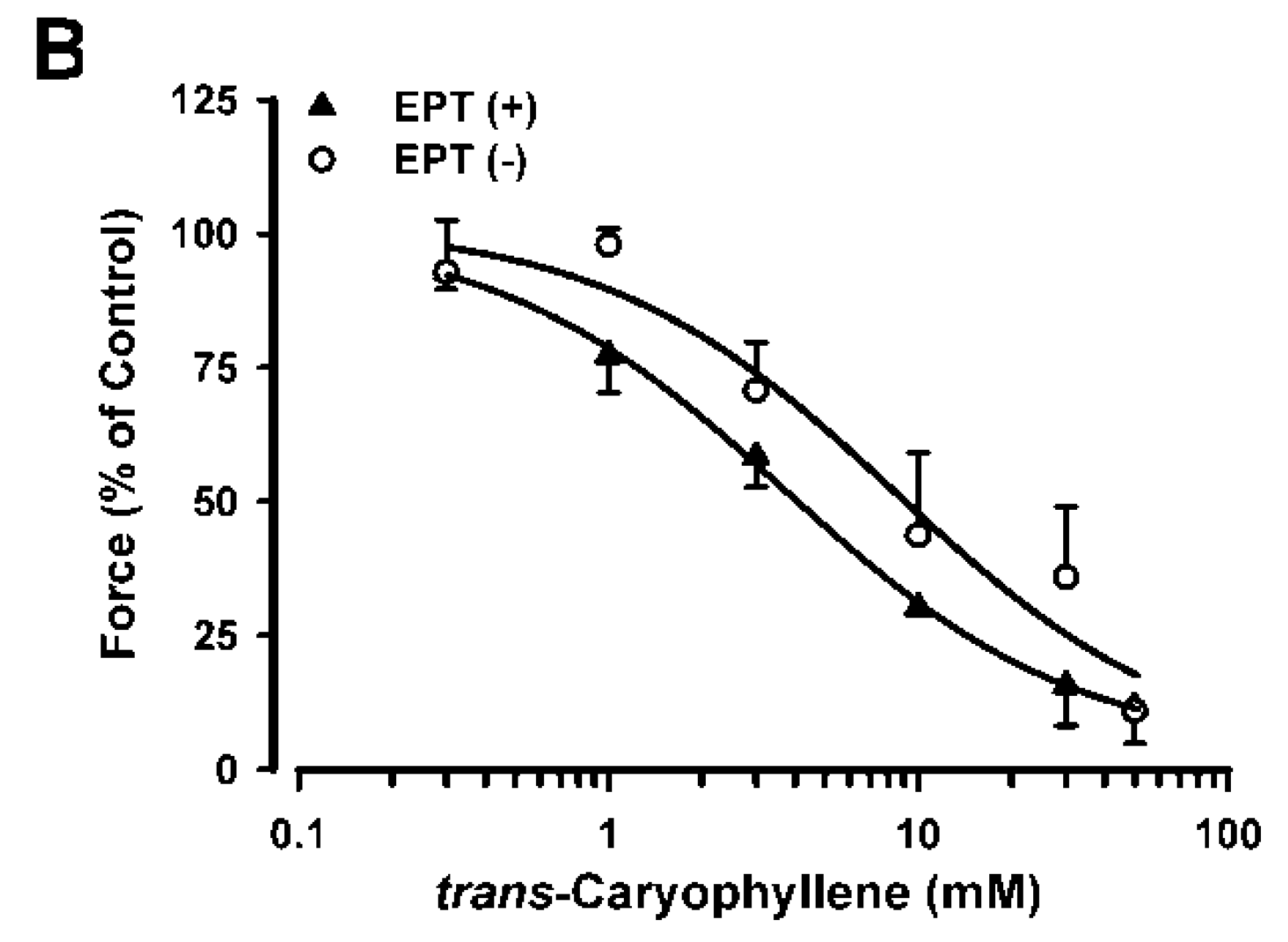
2.2. Inhibitory Effects of trans-Caryophyllene on Contractions Induced by High-K+ in Isolated Rat Trachea with or without Epithelium
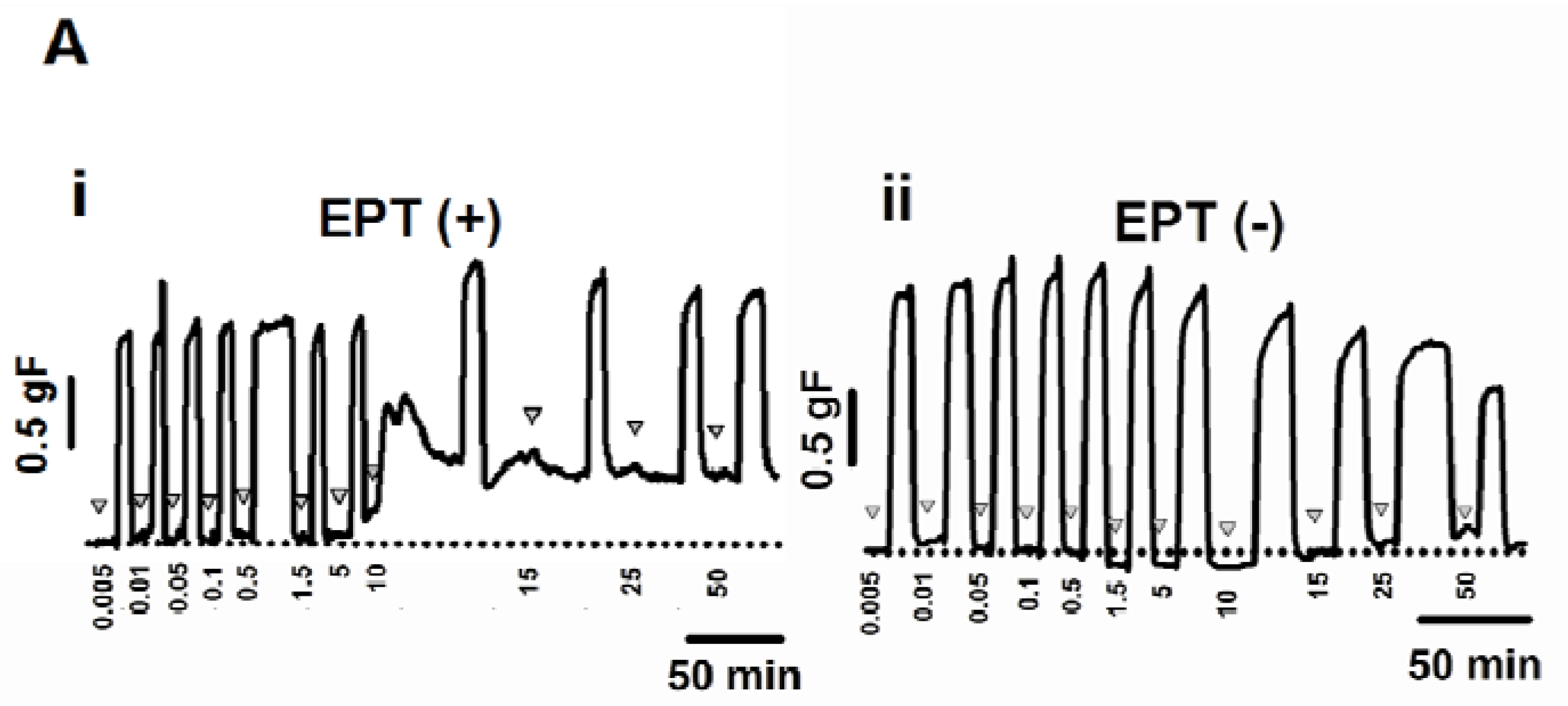
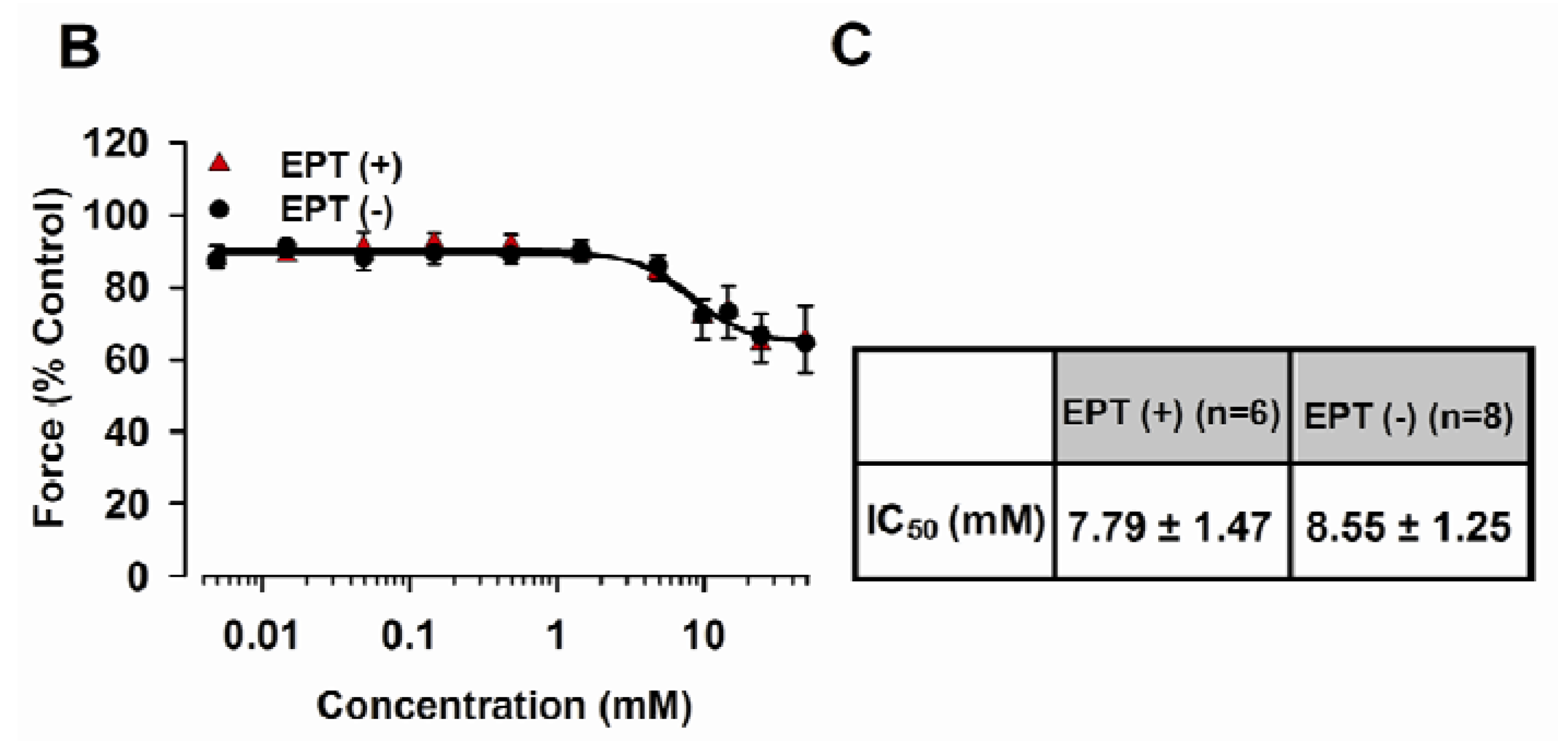
2.3. Inhibitory Effects of trans-Caryophyllene on Contractions Induced by ACh in Isolated Rat Trachea with or without Epithelium
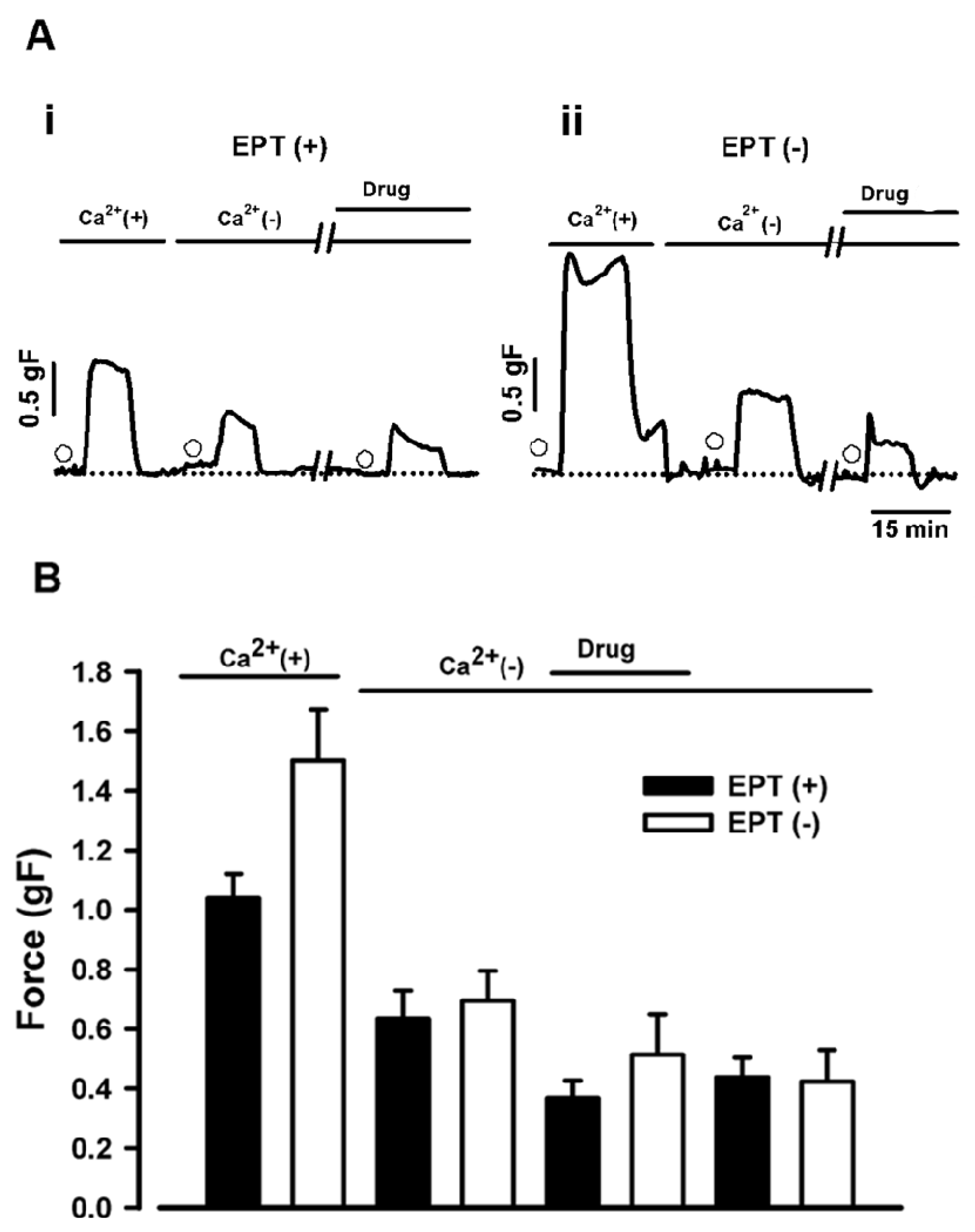
2.4. Effects of trans-Caryophyllene on Contractions Induced by ACh in Isolated Rat Trachea with or without Intact Epithelium in Ca2+-free Solution.
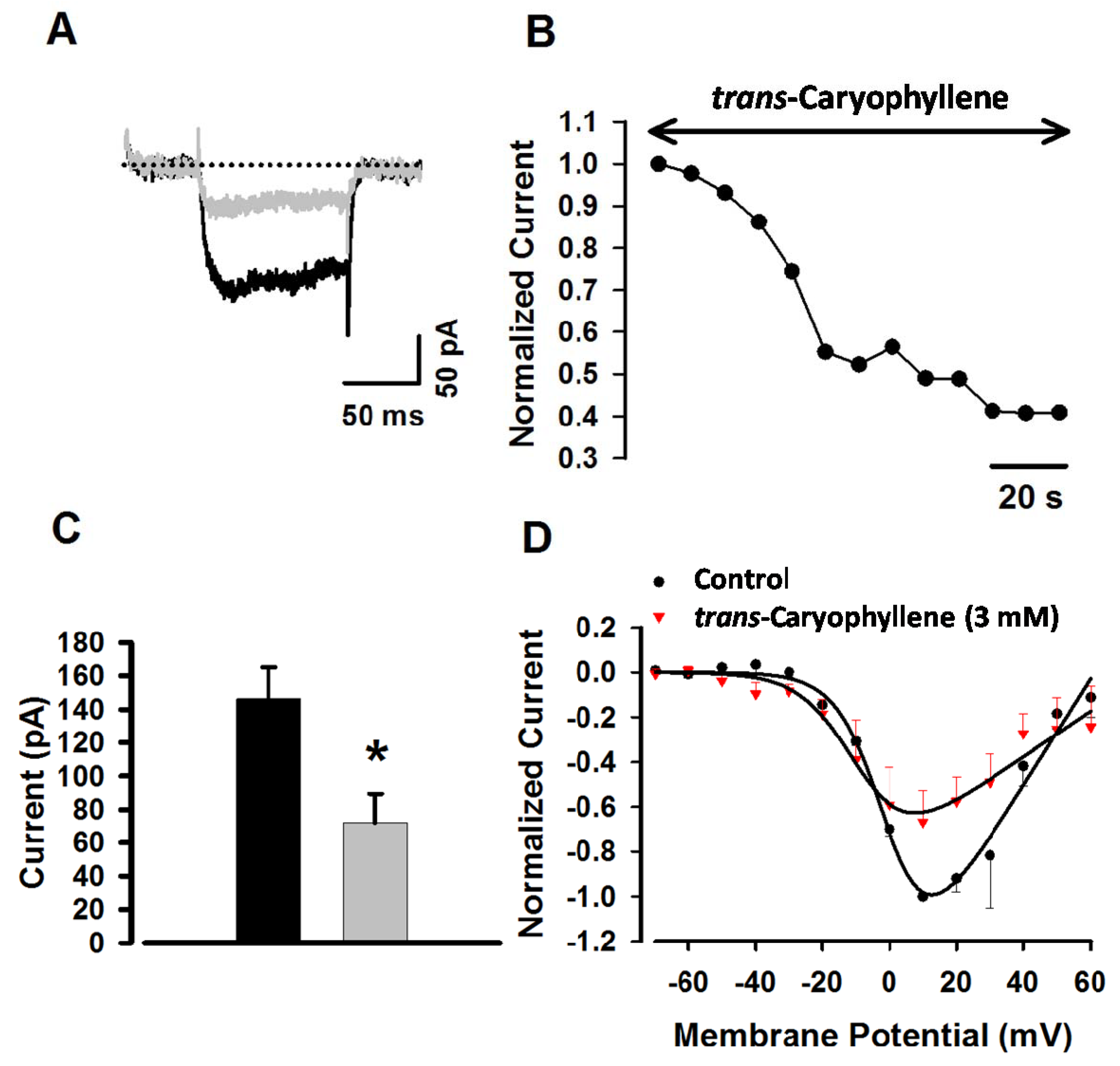
2.5. Effects of trans-Caryophyllene on Isolated Tracheal Smooth Muscle Cells
2.6. Discussion
3. Experimental
3.1. Animals
3.2. Drugs
3.3. Isolated Tissue Preparation
3.3.1. Effects of trans-Caryophyllene on Tracheal Basal Tone
3.3.2. Effects of trans-Caryophyllene on the Contractions Induced by ACh and KCl in Isolated Trachea Maintained in Ca2+-Containing Medium
3.3.3. Effects of trans-Caryophyllene on ACh-Induced Contraction of Rat Isolated Trachea Maintained in Ca2+-Free Medium
3.4. Isolation of Single Tracheal Smooth Muscle Cells
3.5. Membrane Current Recordings
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Craveiro, A.A.; Fernandes, A.G.; Andrade, C.H.S.; Matos, F.J.A.; Alencar, J.W.; Machado, M.I.L. Óleos Essenciais de Plantas do Nordeste; UFC: Fortaleza, Brazil, 1981. [Google Scholar]
- Leal-Cardoso, J.H.; Fonteles, M.C. Pharmacological effects of essential oils of plants of the northeast of Brazil. An. Acad. Bras. Cienc. 1999, 71, 207–213. [Google Scholar] [PubMed]
- Evangelista, G.L.; Coelho-de-Souza, A.N.; Santos, C.F.; Leal-Cardoso, J.H.; Lopes, E.A.; dos Santos, M.V.; Lahlou, S.; Magalhães, P.J. Essential oil of Pterodon polygalaeflorus inhibits electromechanical coupling on rat isolated trachea. J. Ethnopharmacol. 2007, 109, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Astani, A.; Reichling, J.; Schnitzler, P. Screening for Antiviral Activities of Isolated Compounds from Essential Oils. Evid. Based Complement Alternat. Med. 2011. [Google Scholar] [CrossRef] [PubMed]
- Chavan, M.J.; Wakte, P.S.; Shinde, D.B. Analgesic and anti-inflammatory activity of Caryophyllene oxide from Annona squamosa L. bark. Phytomedicine 2010, 17, 149–151. [Google Scholar] [CrossRef] [PubMed]
- Gertsch, J. Anti-inflammatory cannabinoids in diet: Towards a better understanding of CB (2) receptor action? Commun. Integr. Biol. 2008, 1, 26–28. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, E.S.; Passos, G.F.; Medeiros, R.; da Cunha, F.M.; Ferreira, J.; Campos, M.M.; Pianowski, L.F.; Calixto, J.B. Anti-inflammatory effects of compounds alpha-humulene and (−-)-trans-caryophyllene isolated from the essential oil of Cordia verbenacea. Eur. J. Pharmacol. 2007, 569, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, R.; Passos, G.F.; Vitor, C.E.; Koepp, J.; Mazzuco, T.L.; Pianowski, L.F.; Campos, M.M.; Calixto, J.B. Effect of two active compounds obtained from the essential oil of Cordia verbenacea on the acute inflammatory responses elicited by LPS in the rat paw. Br. J. Pharmacol. 2007, 151, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Leonhardt, V.; Leal-Cardoso, J.H.; Lahlou, S.; Albuquerque, A.A.; Porto, R.S.; Celedônio, N.R.; Oliveira, A.C.; Pereira, R.F.; Silva, L.P.; Garcia-Teófilo, T.M.; et al. Antispasmodic effects of essential oil of Pterodon polygalaeflorus and its main constituent beta-caryophyllene on rat isolated ileum. Fundam. Clin. Pharm. 2010, 24, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Janssen, L.J. Ionic mechanisms and Ca (2+) regulation in airway smooth muscle contraction: Do the data contradict dogma? Am. J. Physiol. Lung Cell. Mol. Physiol. 2002, 282, 1161–1178. [Google Scholar] [CrossRef] [PubMed]
- Goldie, R.G.; Fernandes, L.B.; Farmer, S.G.; Hay, D.W. Airway epithelium-derived inhibitory factor. Trends Pharmacol. Sci. 1990, 11, 67–70. [Google Scholar] [CrossRef]
- Grafe, P.; Mayer, C.J.; Wood, J.D. Synaptic modulation of calcium-dependent potassium conductance in myenteric neurones in the guinea-pig. J. Physiol. 1980, 305, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Janssen, L.J.; Hague, C.; Nana, R. Ionic mechanisms underlying electrical slow waves in canine airway smooth muscle. Am. J. Physiol. 1998, 275, 516–523. [Google Scholar] [CrossRef]
- Hille, B. Ion Channels of Excitable Membranes, 3rd ed.; Sinauer Associates: Sunderland, UK, 2001. [Google Scholar]
- Murray, R.K.; Kotlikoff, M.I. Receptor-activated calcium influx in human airway smooth cells. J. Physiol. 1991, 435, 123–144. [Google Scholar] [CrossRef] [PubMed]
- Somlyo, A.V.; Bond, M.; Somlyo, A.P.; Scarpa, A. Inositol trisphosphate-induced calcium release and contraction in vascular smooth muscle. Proc. Natl. Acad. Sci. USA 1985, 82, 5231–5235. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, D.; Rabe, K.F. The role of leukotrienes in the regulation of tone and responsiveness in isolated human airways. Am. J. Resp. Crit. Care. Med. 2000, 161, S62–S67. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; McMahon, T.J.; Zhang, Z.S.; Stiber, J.A.; Meissner, G.; Eu, J.P. Excitation-contraction coupling in airway smooth muscle. J. Biol. Chem. 2006, 281, 31143–31151. [Google Scholar] [CrossRef] [PubMed]
- Coburn, R.F.; Baron, C.B. Coupling mechanisms in airway smooth muscle. Am. J. Physiol. Lung. Cell. Mol. Physiol. 1990, 258, L119–L133. [Google Scholar] [CrossRef] [PubMed]
- Cuthbert, N.J.; Gardiner, P.J.; Nash, K.; Poll, C.T. Roles of Ca2+ influx and Ca2+ release in agonist-induced contractions in guinea-pig trachea. Am. J. Physiol. Lung. Cell. Mol. Physiol. 1994, 266, L620–L627. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, B.G.; Silva, A.S.B.; Souza, G.E.P.; Figueiredo, J.G.; Cunha, F.Q.; Lahlou, S.; da Silva, J.K.R.; Maia, J.G.S.; Sousa, P.J.C. Chemical composition, antinociceptive and anti-inflammatory effects in rodents of the essential oil of Peperomia serpens (Sw.) Loud. J. Ethnopharm. 2011, 138, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y. Recent Advances in Mechanisms and Treatments of Airway Remodeling in Asthma: A Message from the Bench Side to the Clinic. Korean J. Int. Med. 2011, 26, 367–383. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |







© 2012 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pinho-da-Silva, L.; Mendes-Maia, P.V.; Teófilo, T.M.D.N.G.; Barbosa, R.; Ceccatto, V.M.; Coelho-de-Souza, A.N.; Santos Cruz, J.; Leal-Cardoso, J.H. trans-Caryophyllene, a Natural Sesquiterpene, Causes Tracheal Smooth Muscle Relaxation through Blockade of Voltage-Dependent Ca2+ Channels. Molecules 2012, 17, 11965-11977. https://doi.org/10.3390/molecules171011965
Pinho-da-Silva L, Mendes-Maia PV, Teófilo TMDNG, Barbosa R, Ceccatto VM, Coelho-de-Souza AN, Santos Cruz J, Leal-Cardoso JH. trans-Caryophyllene, a Natural Sesquiterpene, Causes Tracheal Smooth Muscle Relaxation through Blockade of Voltage-Dependent Ca2+ Channels. Molecules. 2012; 17(10):11965-11977. https://doi.org/10.3390/molecules171011965
Chicago/Turabian StylePinho-da-Silva, Leidiane, Paula Virgínia Mendes-Maia, Taylena Maria Do Nascimento Garcia Teófilo, Roseli Barbosa, Vania Marilande Ceccatto, Andrelina Noronha Coelho-de-Souza, Jader Santos Cruz, and José Henrique Leal-Cardoso. 2012. "trans-Caryophyllene, a Natural Sesquiterpene, Causes Tracheal Smooth Muscle Relaxation through Blockade of Voltage-Dependent Ca2+ Channels" Molecules 17, no. 10: 11965-11977. https://doi.org/10.3390/molecules171011965