Proteomic and Metabolomic Correlates of Healthy Dietary Patterns: The Framingham Heart Study

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sample
2.2. Protein Quantification
2.3. Metabolite Quantification
2.4. Dietary Assessment
2.5. Dietary Pattern Indices
2.6. Covariate Assessment
2.7. Statistical Analysis
2.8. Enrichment Analysis
Overlap between Dietary Protein and Metabolite Quantitative Trait Loci and Prior Genome-Wide Association Study Risk Loci
3. Results
3.1. Sample Characteristics
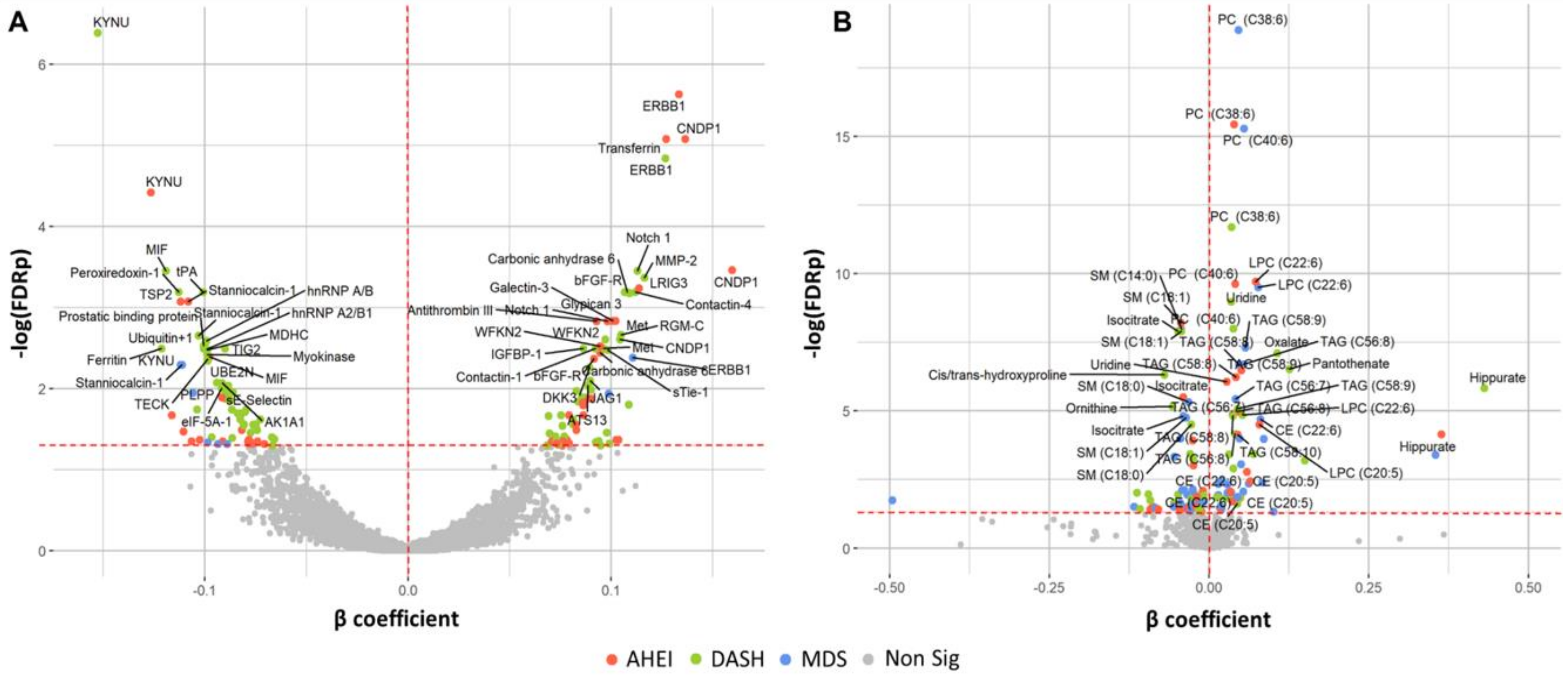
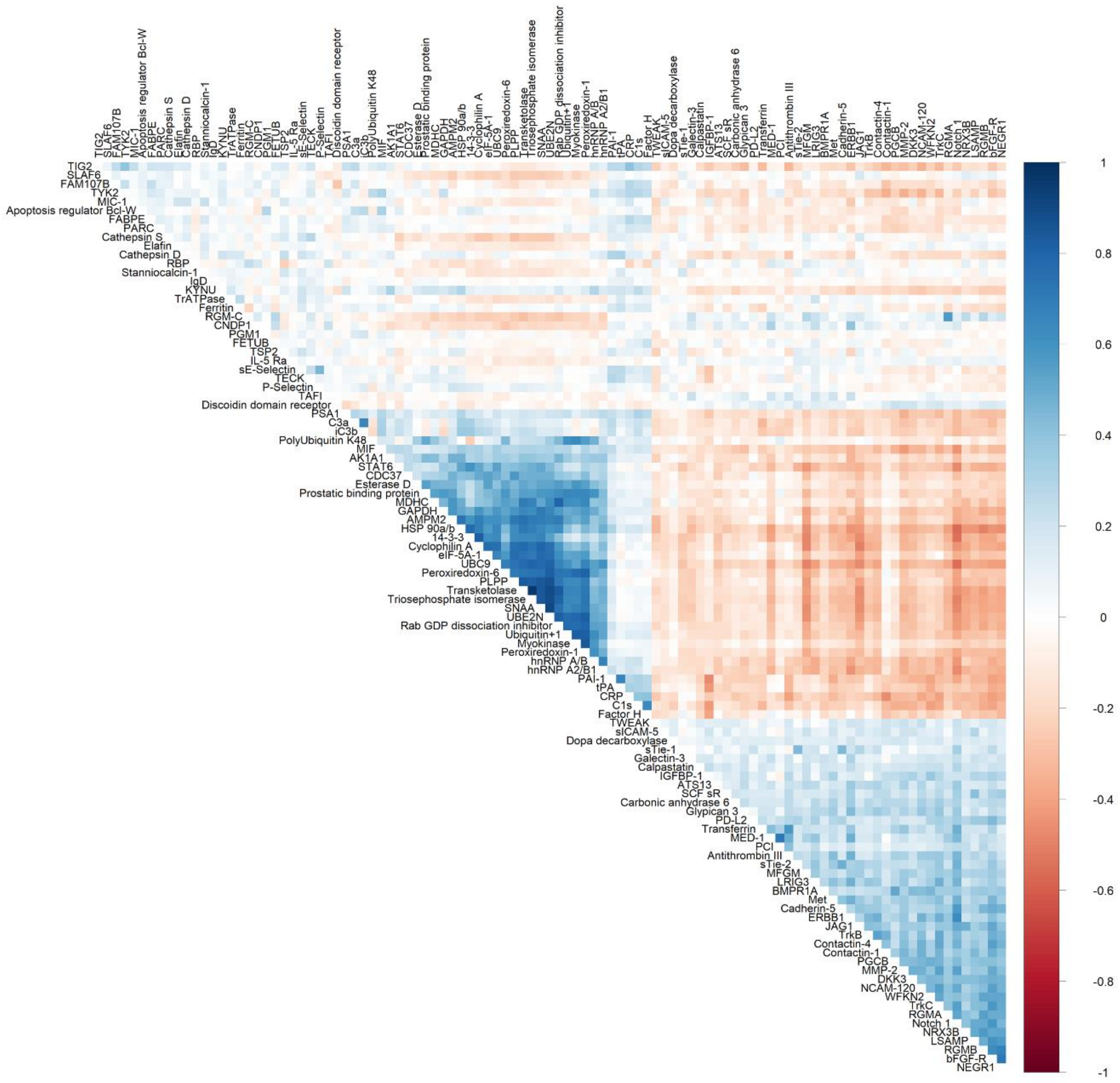
3.2. Associations of Dietary Patterns with Plasma Protein Concentrations
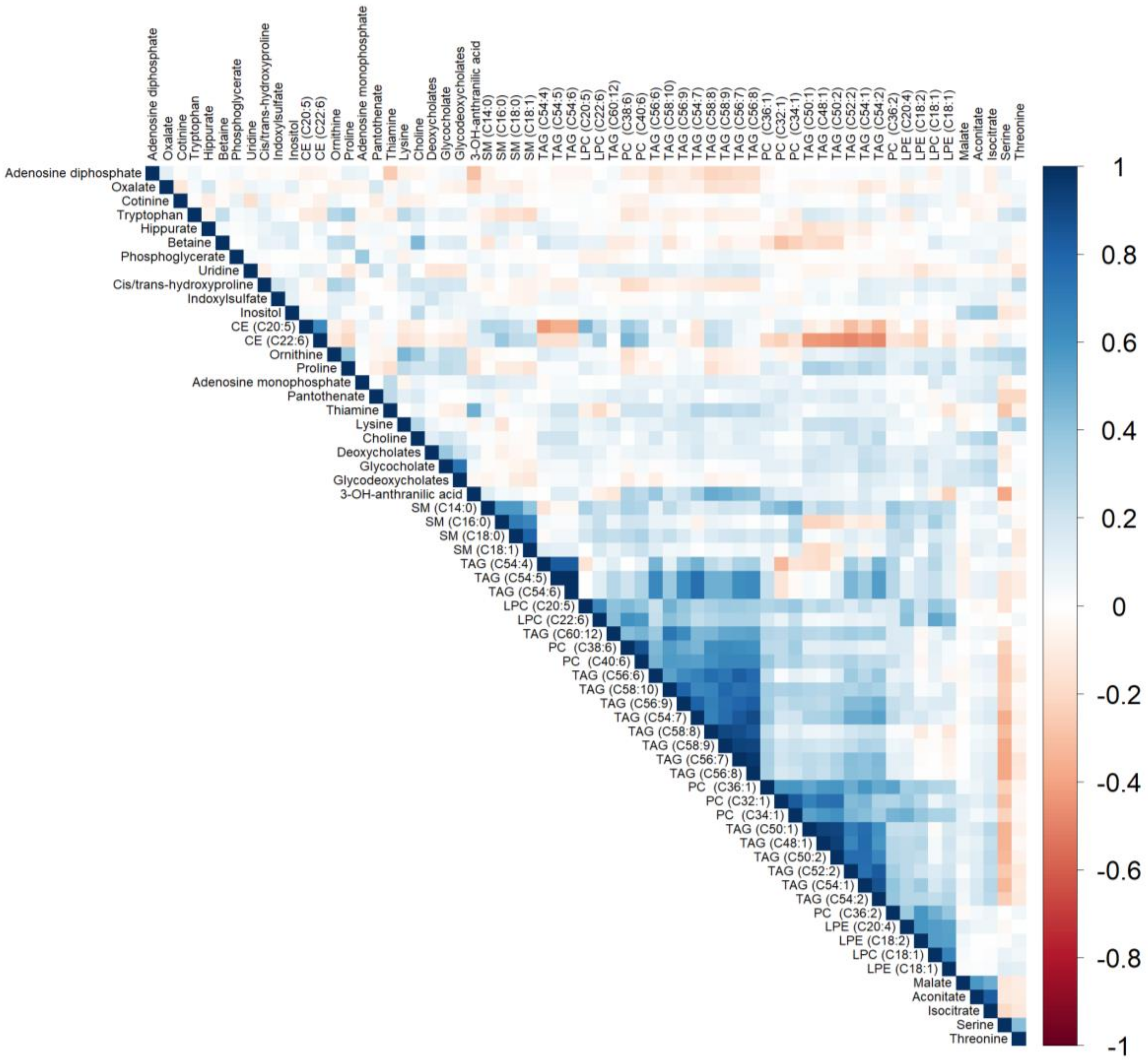
3.3. Associations of Dietary Patterns with Plasma Metabolite Concentrations
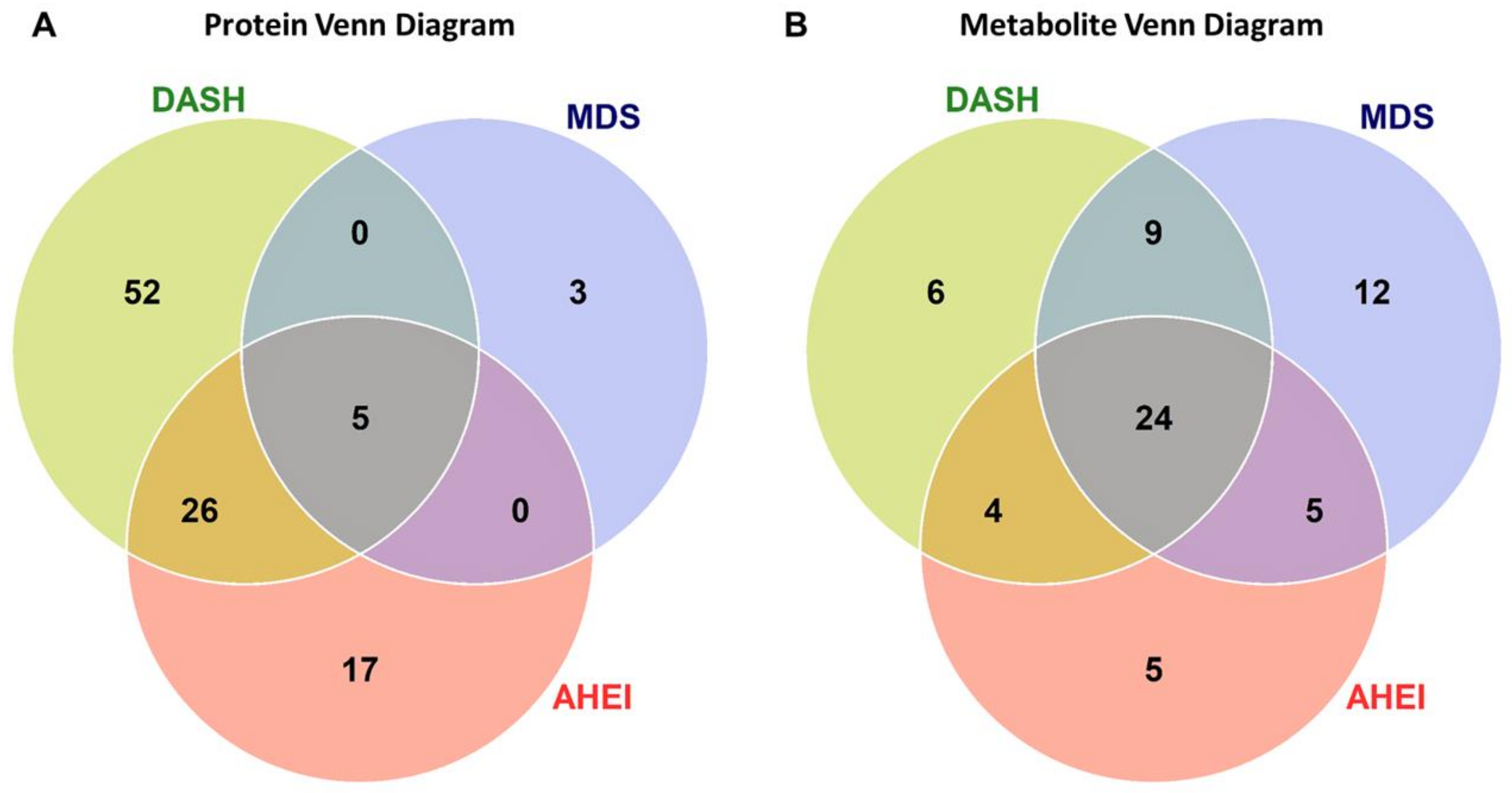
3.4. Protein Enrichment Analysis
3.5. Metabolite Enrichment
3.6. Overlap between Dietary Protein and Metabolite Quantitative Trait Loci and Prior Genome-Wide Association Study Risk Loci
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Micha, R.; Peñalvo, J.L.; Cudhea, F.; Imamura, F.; Rehm, C.D.; Mozaffarian, D. Association Between Dietary Factors and Mortality From Heart Disease, Stroke, and Type 2 Diabetes in the United States. JAMA 2017, 317, 912–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.-I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet Supplemented with Extra-Virgin Olive Oil or Nuts. N. Engl. J. Med. 2018, 378, e34. [Google Scholar] [CrossRef] [PubMed]
- Appel, L.J.; Moore, T.J.; Obarzanek, E.; Vollmer, W.M.; Svetkey, L.P.; Sacks, F.M.; Bray, G.A.; Vogt, T.M.; Cutler, J.A.; Windhauser, M.M.; et al. A Clinical Trial of the Effects of Dietary Patterns on Blood Pressure. N. Engl. J. Med. 1997, 336, 1117–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reedy, J.; Krebs-Smith, S.M.; Miller, P.E.; Liese, A.D.; Kahle, L.L.; Park, Y.; Subar, A.F. Higher diet quality is associated with decreased risk of all-cause, cardiovascular disease, and cancer mortality among older adults. J. Nutr. 2014, 144, 881–889. [Google Scholar] [CrossRef] [Green Version]
- Harmon, B.E.; Boushey, C.J.; Shvetsov, Y.B.; Ettienne, R.; Reedy, J.; Wilkens, L.R.; Le Marchand, L.; Henderson, B.E.; Kolonel, L.N. Associations of key diet-quality indexes with mortality in the Multiethnic Cohort: The Dietary Patterns Methods Project. Am. J. Clin. Nutr. 2015, 101, 587–597. [Google Scholar] [CrossRef] [Green Version]
- Brennan, L.; Hu, F.B. Metabolomics-Based Dietary Biomarkers in Nutritional Epidemiology-Current Status and Future Opportunities. Mol. Nutr. Food Res. 2019, 63, e1701064. [Google Scholar] [CrossRef]
- Playdon, M.C.; Ziegler, R.G.; Sampson, J.N.; Stolzenberg-Solomon, R.; Thompson, H.J.; Irwin, M.L.; Mayne, S.T.; Hoover, R.N.; Moore, S.C. Nutritional metabolomics and breast cancer risk in a prospective study. Am. J. Clin. Nutr. 2017, 106, 637–649. [Google Scholar] [CrossRef]
- Guertin, K.A.; Loftfield, E.; Boca, S.M.; Sampson, J.N.; Moore, S.C.; Xiao, Q.; Huang, W.-Y.; Xiong, X.; Freedman, N.D.; Cross, A.J.; et al. Serum biomarkers of habitual coffee consumption may provide insight into the mechanism underlying the association between coffee consumption and colorectal cancer. Am. J. Clin. Nutr. 2015, 101, 1000–1011. [Google Scholar] [CrossRef]
- Akbaraly, T.; Würtz, P.; Singh-Manoux, A.; Shipley, M.J.; Haapakoski, R.; Lehto, M.; Desrumaux, C.; Kähönen, M.; Lehtimäki, T.; Mikkilä, V.; et al. Association of circulating metabolites with healthy diet and risk of cardiovascular disease: Analysis of two cohort studies. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Kannel, W.B.; Feinleib, M.; McNamara, P.M.; Garrison, R.J.; Castelli, W.P. An investigation of coronary heart disease in families. The Framingham offspring study. Am. J. Epidemiol. 1979, 110, 281–290. [Google Scholar] [CrossRef]
- Ngo, D.; Sinha, S.; Shen, D.; Kuhn, E.W.; Keyes, M.J.; Shi, X.; Benson, M.D.; O’Sullivan, J.F.; Keshishian, H.; Farrell, L.A.; et al. Aptamer-Based Proteomic Profiling Reveals Novel Candidate Biomarkers and Pathways in Cardiovascular Disease. Circulation 2016, 134, 270–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, D.; Benson, M.D.; Ngo, D.; Yang, Q.; Larson, M.G.; Wang, T.J.; Trinquart, L.; McManus, D.D.; Lubitz, S.A.; Ellinor, P.T.; et al. Proteomics Profiling and Risk of New-Onset Atrial Fibrillation: Framingham Heart Study. J. Am. Heart Assoc. 2019, 8, e010976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite profiles and the risk of developing diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Rhee, E.P.; Cheng, S.; Larson, M.G.; Walford, G.A.; Lewis, G.D.; McCabe, E.; Yang, E.; Farrell, L.; Fox, C.S.; O’Donnell, C.J.; et al. Lipid profiling identifies a triacylglycerol signature of insulin resistance and improves diabetes prediction in humans. J. Clin. Investig. 2011, 121, 1402–1411. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.J.; Ngo, D.; Psychogios, N.; Dejam, A.; Larson, M.G.; Vasan, R.S.; Ghorbani, A.; O’Sullivan, J.; Cheng, S.; Rhee, E.P.; et al. 2-Aminoadipic acid is a biomarker for diabetes risk. J. Clin. Investig. 2013, 123, 4309–4317. [Google Scholar] [CrossRef]
- Feskanich, D.; Rimm, E.B.; Giovannucci, E.L.; Colditz, G.A.; Stampfer, M.J.; Litin, L.B.; Willett, W.C. Reproducibility and validity of food intake measurements from a semiquantitative food frequency questionnaire. J. Am. Diet. Assoc. 1993, 93, 790–796. [Google Scholar] [CrossRef]
- Willett, W. Nutritional Epidemiology; Oxford University Press: Oxford, UK, 2012; ISBN 978-0-19-024084-4. [Google Scholar]
- Fung, T.T.; Rexrode, K.M.; Mantzoros, C.S.; Manson, J.E.; Willett, W.C.; Hu, F.B. Mediterranean Diet and Incidence of and Mortality From Coronary Heart Disease and Stroke in Women. Circulation 2009, 119, 1093–1100. [Google Scholar] [CrossRef] [Green Version]
- Fung, T.T.; Chiuve, S.E.; McCullough, M.L.; Rexrode, K.M.; Logroscino, G.; Hu, F.B. Adherence to a DASH-style diet and risk of coronary heart disease and stroke in women. Arch. Intern. Med. 2008, 168, 713–720. [Google Scholar] [CrossRef] [Green Version]
- Chiuve, S.E.; Fung, T.T.; Rimm, E.B.; Hu, F.B.; McCullough, M.L.; Wang, M.; Stampfer, M.J.; Willett, W.C. Alternative dietary indices both strongly predict risk of chronic disease. J. Nutr. 2012, 142, 1009–1018. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Hennein, R.; Liu, C.; Long, M.T.; Hoffmann, U.; Jacques, P.F.; Lichtenstein, A.H.; Hu, F.B.; Levy, D. Improved Diet Quality Associates With Reduction in Liver Fat, Particularly in Individuals With High Genetic Risk Scores for Nonalcoholic Fatty Liver Disease. Gastroenterology 2018, 155, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Kannel, W.B.; Sorlie, P. Some health benefits of physical activity. The Framingham Study. Arch. Intern. Med. 1979, 139, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. WebGestalt 2019: Gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Rhee, E.P.; Ho, J.E.; Chen, M.-H.; Shen, D.; Cheng, S.; Larson, M.G.; Ghorbani, A.; Shi, X.; Helenius, I.T.; O’Donnell, C.J.; et al. A Genome-Wide Association Study of the Human Metabolome in a Community-Based Cohort. Cell Metab. 2013, 18, 130–143. [Google Scholar] [CrossRef] [Green Version]
- Benson, M.D.; Yang, Q.; Ngo, D.; Zhu, Y.; Shen, D.; Farrell, L.A.; Sinha, S.; Keyes, M.J.; Vasan, R.S.; Larson, M.G.; et al. Genetic Architecture of the Cardiovascular Risk Proteome. Circulation 2018, 137, 1158–1172. [Google Scholar] [CrossRef]
- MacArthur, J.; Bowler, E.; Cerezo, M.; Gil, L.; Hall, P.; Hastings, E.; Junkins, H.; McMahon, A.; Milano, A.; Morales, J.; et al. The new NHGRI-EBI Catalog of published genome-wide association studies (GWAS Catalog). Nucleic Acids Res. 2017, 45, D896–D901. [Google Scholar] [CrossRef]
- Van der Harst, P.; Verweij, N. Identification of 64 Novel Genetic Loci Provides an Expanded View on the Genetic Architecture of Coronary Artery Disease. Circ. Res. 2018, 122, 433–443. [Google Scholar] [CrossRef]
- Lindström, S.; Wang, L.; Smith, E.N.; Gordon, W.; van Hylckama Vlieg, A.; de Andrade, M.; Brody, J.A.; Pattee, J.W.; Haessler, J.; Brumpton, B.M.; et al. Genomic and transcriptomic association studies identify 16 novel susceptibility loci for venous thromboembolism. Blood 2019, 134, 1645–1657. [Google Scholar] [CrossRef]
- Lemaitre, R.N.; Tanaka, T.; Tang, W.; Manichaikul, A.; Foy, M.; Kabagambe, E.K.; Nettleton, J.A.; King, I.B.; Weng, L.-C.; Bhattacharya, S.; et al. Genetic loci associated with plasma phospholipid n-3 fatty acids: A meta-analysis of genome-wide association studies from the CHARGE Consortium. PLoS Genet. 2011, 7, e1002193. [Google Scholar] [CrossRef] [Green Version]
- Guan, W.; Steffen, B.T.; Lemaitre, R.N.; Wu, J.H.Y.; Tanaka, T.; Manichaikul, A.; Foy, M.; Rich, S.S.; Wang, L.; Nettleton, J.A.; et al. Genome-wide association study of plasma N6 polyunsaturated fatty acids within the cohorts for heart and aging research in genomic epidemiology consortium. Circ. Cardiovasc. Genet. 2014, 7, 321–331. [Google Scholar] [CrossRef] [Green Version]
- Dupuis, J.; Langenberg, C.; Prokopenko, I.; Saxena, R.; Soranzo, N.; Jackson, A.U.; Wheeler, E.; Glazer, N.L.; Bouatia-Naji, N.; Gloyn, A.L.; et al. New genetic loci implicated in fasting glucose homeostasis and their impact on type 2 diabetes risk. Nat. Genet. 2010, 42, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Wojcik, G.L.; Graff, M.; Nishimura, K.K.; Tao, R.; Haessler, J.; Gignoux, C.R.; Highland, H.M.; Patel, Y.M.; Sorokin, E.P.; Avery, C.L.; et al. Genetic analyses of diverse populations improves discovery for complex traits. Nature 2019, 570, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.P.; Goel, A.; Butterworth, A.S.; Kanoni, S.; Webb, T.R.; Marouli, E.; Zeng, L.; Ntalla, I.; Lai, F.Y.; Hopewell, J.C.; et al. Association analyses based on false discovery rate implicate new loci for coronary artery disease. Nat. Genet. 2017, 49, 1385–1391. [Google Scholar] [CrossRef] [PubMed]
- Gold, L.; Ayers, D.; Bertino, J.; Bock, C.; Bock, A.; Brody, E.N.; Carter, J.; Dalby, A.B.; Eaton, B.E.; Fitzwater, T.; et al. Aptamer-based multiplexed proteomic technology for biomarker discovery. PLoS ONE 2010, 5, e15004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carayol, J.; Chabert, C.; Cara, A.D.; Armenise, C.; Lefebvre, G.; Langin, D.; Viguerie, N.; Metairon, S.; Saris, W.H.M.; Astrup, A.; et al. Protein quantitative trait locus study in obesity during weight-loss identifies a leptin regulator. Nat. Commun. 2017, 8, 1–14. [Google Scholar] [CrossRef]
- Ruffieux, H.; Carayol, J.; Harper, M.E.; Dent, R.; Saris, W.H.M.; Astrup, A.; Davison, A.C.; Hager, J.; Valsesia, A. A large-scale multivariate pQTL study sheds light on the genetic architecture of obesity. bioRxiv 2019, 524405. [Google Scholar] [CrossRef] [Green Version]
- García-Bailo, B.; Brenner, D.R.; Nielsen, D.; Lee, H.-J.; Domanski, D.; Kuzyk, M.; Borchers, C.H.; Badawi, A.; Karmali, M.A.; El-Sohemy, A. Dietary patterns and ethnicity are associated with distinct plasma proteomic groups. Am. J. Clin. Nutr. 2012, 95, 352–361. [Google Scholar] [CrossRef]
- Warensjö Lemming, E.; Byberg, L.; Stattin, K.; Ahmad, S.; Lind, L.; Elmståhl, S.; Larsson, S.C.; Wolk, A.; Michaëlsson, K. Dietary Pattern Specific Protein Biomarkers for Cardiovascular Disease: A Cross-Sectional Study in 2 Independent Cohorts. J. Am. Heart Assoc. 2019, 8, e011860. [Google Scholar] [CrossRef]
- Wang, L.; Huang, Z.; Huang, W.; Chen, X.; Shan, P.; Zhong, P.; Khan, Z.; Wang, J.; Fang, Q.; Liang, G.; et al. Inhibition of epidermal growth factor receptor attenuates atherosclerosis via decreasing inflammation and oxidative stress. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Simons, K.H.; de Jong, A.; Jukema, J.W.; de Vries, M.R.; Arens, R.; Quax, P.H.A. T cell co-stimulation and co-inhibition in cardiovascular disease: A double-edged sword. Nat. Rev. Cardiol. 2019, 16, 325–343. [Google Scholar] [CrossRef]
- Nai, W.; Threapleton, D.; Lu, J.; Zhang, K.; Wu, H.; Fu, Y.; Wang, Y.; Ou, Z.; Shan, L.; Ding, Y.; et al. Identification of novel genes and pathways in carotid atheroma using integrated bioinformatic methods. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, M.S.; Heald, A.H.; Gibson, J.M.; Cruickshank, J.K.; Dunger, D.B.; Wareham, N.J. Circulating concentrations of insulin-like growth factor-I and development of glucose intolerance: A prospective observational study. Lancet 2002, 359, 1740–1745. [Google Scholar] [CrossRef]
- Lewitt, M.S.; Hilding, A.; Ostenson, C.-G.; Efendic, S.; Brismar, K.; Hall, K. Insulin-like growth factor-binding protein-1 in the prediction and development of type 2 diabetes in middle-aged Swedish men. Diabetologia 2008, 51, 1135–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho Jennifer, E.; Lyass, A.; Courchesne, P.; Chen, G.; Liu, C.; Yin, X.; Hwang, S.-J.; Massaro, J.M.; Larson, M.G.; Levy, D. Protein Biomarkers of Cardiovascular Disease and Mortality in the Community. J. Am. Heart Assoc. 2018, 7, e008108. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.E.; Liu, C.; Lyass, A.; Courchesne, P.; Pencina, M.J.; Vasan, R.S.; Larson, M.G.; Levy, D. Galectin-3, a Marker of Cardiac Fibrosis, Predicts Incident Heart Failure in the Community. J. Am. Coll. Cardiol. 2012, 60, 1249–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forman, J.P.; Stampfer, M.J.; Curhan, G.C. Diet and lifestyle risk factors associated with incident hypertension in women. JAMA 2009, 302, 401–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwingshackl, L.; Hoffmann, G. Diet quality as assessed by the Healthy Eating Index, the Alternate Healthy Eating Index, the Dietary Approaches to Stop Hypertension score, and health outcomes: A systematic review and meta-analysis of cohort studies. J. Acad. Nutr. Diet. 2015, 115, 780–800.e5. [Google Scholar] [CrossRef]
- Playdon, M.C.; Moore, S.C.; Derkach, A.; Reedy, J.; Subar, A.F.; Sampson, J.N.; Albanes, D.; Gu, F.; Kontto, J.; Lassale, C.; et al. Identifying biomarkers of dietary patterns by using metabolomics. Am. J. Clin. Nutr. 2017, 105, 450–465. [Google Scholar] [CrossRef] [Green Version]
- McCullough, M.L.; Maliniak, M.L.; Stevens, V.L.; Carter, B.D.; Hodge, R.A.; Wang, Y. Metabolomic markers of healthy dietary patterns in US postmenopausal women. Am. J. Clin. Nutr. 2019, 109, 1439–1451. [Google Scholar] [CrossRef]
- Rebholz, C.M.; Lichtenstein, A.H.; Zheng, Z.; Appel, L.J.; Coresh, J. Serum untargeted metabolomic profile of the Dietary Approaches to Stop Hypertension (DASH) dietary pattern. Am. J. Clin. Nutr. 2018, 108, 243–255. [Google Scholar] [CrossRef]
- Waterworth, D.M.; Ricketts, S.L.; Song, K.; Chen, L.; Zhao, J.H.; Ripatti, S.; Aulchenko, Y.S.; Zhang, W.; Yuan, X.; Lim, N.; et al. Genetic variants influencing circulating lipid levels and risk of coronary artery disease. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2264–2276. [Google Scholar] [CrossRef]
- Hoffmann, T.J.; Theusch, E.; Haldar, T.; Ranatunga, D.K.; Jorgenson, E.; Medina, M.W.; Kvale, M.N.; Kwok, P.-Y.; Schaefer, C.; Krauss, R.M.; et al. A large electronic health record-based genome-wide study of serum lipids. Nat. Genet. 2018, 50, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Stegemann, C.; Pechlaner, R.; Willeit, P.; Langley, S.R.; Mangino, M.; Mayr, U.; Menni, C.; Moayyeri, A.; Santer, P.; Rungger, G.; et al. Lipidomics Profiling and Risk of Cardiovascular Disease in the Prospective Population-Based Bruneck Study. Circulation 2014, 129, 1821–1831. [Google Scholar] [CrossRef] [Green Version]
- Rhee, E.P.; Yang, Q.; Yu, B.; Liu, X.; Cheng, S.; Deik, A.; Pierce, K.A.; Bullock, K.; Ho, J.E.; Levy, D.; et al. An exome array study of the plasma metabolome. Nat. Commun. 2016, 7, 12360. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.D.; Zheng, Y.; Toledo, E.; Razquin, C.; Ruiz-Canela, M.; Guasch-Ferré, M.; Yu, E.; Corella, D.; Gómez-Gracia, E.; Fiol, M.; et al. Lipid metabolic networks, Mediterranean diet and cardiovascular disease in the PREDIMED trial. Int. J. Epidemiol. 2018, 47, 1830–1845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toledo, E.; Wang, D.D.; Ruiz-Canela, M.; Clish, C.B.; Razquin, C.; Zheng, Y.; Guasch-Ferré, M.; Hruby, A.; Corella, D.; Gómez-Gracia, E.; et al. Plasma lipidomic profiles and cardiovascular events in a randomized intervention trial with the Mediterranean diet. Am. J. Clin. Nutr. 2017, 106, 973–983. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B.; Bajaj, J.S. Bile Acids and the Gut Microbiome. Curr. Opin. Gastroenterol. 2014, 30, 332–338. [Google Scholar] [CrossRef] [Green Version]
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.-Y.; Tarng, D.-C. Diet, gut microbiome and indoxyl sulphate in chronic kidney disease patients. Nephrology (Carlton) 2018, 23, 16–20. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.H.W.; Wang, Z.; Levison, B.S.; Koeth, R.A.; Britt, E.B.; Fu, X.; Wu, Y.; Hazen, S.L. Intestinal Microbial Metabolism of Phosphatidylcholine and Cardiovascular Risk. N. Engl. J. Med. 2013, 368, 1575–1584. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Men | Women | |
|---|---|---|
| n = 2208 | Mean ± SD | Mean ± SD |
| n = 1054 | n = 1154 | |
| Age, yr | 55.7 ± 9.9 | 54.5 ± 9.6 |
| Current smoker, n (%) | 190 (18%) | 210 (18%) |
| Physical activity score | 36.0 ± 7.4 | 33.4 ± 4.7 |
| BMI, kg/m2 | 28.2 ± 4.1 | 26.7 ± 5.4 |
| Lipid-lowering medications, n (%) | 101 (10) | 67 (6) |
| Anti-hypertensive medication, n (%) | 238 (23) | 195 (17) |
| Energy intake, kcal/day | 1992 ± 642 | 1742 ± 566 |
| AHEI (0–110) | 48.0 ± 9.6 | 51.7 ± 9.7 |
| DASH (0–40) | 23.0 ± 4.9 | 25.3 ± 4.7 |
| MDS (0–25) | 12.3 ± 4.3 | 12.0 ± 4.4 |
| Food Group Intake | ||
| Vegetables, svg/d | 2.0 ± 1.3 | 2.4 ± 1.7 |
| Fruits, svg/d | 1.2 ± 1.0 | 1.4 ± 1.2 |
| Low-fat dairy, svg/d | 0.8 ± 0.9 | 0.9 ± 1.0 |
| Nuts, svg/wk | 0.4 ± 0.6 | 0.2 ± 0.4 |
| Legumes, svg/wk | 0.3 ± 0.3 | 0.3 ± 0.3 |
| Whole grains, svg/wk | 1.2 ± 1.2 | 1.2 ± 1.2 |
| Red/processed meat, svg/wk | 0.9 ± 0.7 | 0.6 ± 0.5 |
| Fish, svg/wk | 0.3 ± 0.3 | 0.3 ± 0.3 |
| Sugar-sweetened beverage & fruit juice, svg/wk | 1.4 ± 1.2 | 1.1 ± 1.2 |
| Sugar-sweetened beverage, svg/wk | 0.5 ± 0.8 | 0.3 ± 0.6 |
| Fruit juice, svg/wk | 0.9 ± 0.9 | 0.8 ± 0.9 |
| Alcohol, gm/day | 15.4 ± 20.0 | 6.7 ± 11.2 |
| MUFA:SFA | 1.1 ± 0.2 | 1.1 ± 0.2 |
| MUFA, %kcal | 11.3 ± 2.7 | 10.9 ± 2.6 |
| SFA, % kcal | 10.5 ± 3.0 | 10.3 ± 2.8 |
| PUFA, % kcal | 5.7 ± 1.7 | 6.0 ± 1.7 |
| Trans fat, % kcal | 1.6 ± 0.8 | 1.4 ± 0.7 |
| Omega-3 fatty acids, mg/day | 257.8 ± 235.5 | 253.2 ± 217.8 |
| Name | Type/Class | Diet 2 | AHEI | DASH | MDS | |||
|---|---|---|---|---|---|---|---|---|
| Β 3 | SE | β | SE | β | SE | |||
| Carbonic anhydrase 6 | Enzyme | DASH | 0.095 | 0.023 | 0.108 * | 0.023 | 0.085 | 0.025 |
| Carnosine dipeptidase 1 (CNDP1) | Peptidase | AHEI, DASH | 0.135 * | 0.024 | 0.105 * | 0.025 | 0.085 | 0.026 |
| Contactin 4 | Enzyme | DASH | 0.051 | 0.024 | 0.112 * | 0.024 | 0.075 | 0.025 |
| Epidermal growth factor receptor (ERBB1) | Kinase | AHEI, MDS, DASH | 0.133 * | 0.022 | 0.127 * | 0.023 | 0.111 * | 0.024 |
| Fibroblast growth factor receptor 1 (bFGF-R) | Kinase | AHEI,DASH | 0.092 * | 0.023 | 0.107 * | 0.023 | 0.076 | 0.024 |
| Galectin 3 | Other | AHEI | 0.101 * | 0.023 | 0.073 | 0.024 | 0.076 | 0.025 |
| Glypican 3 | Other | DASH | 0.057 | 0.024 | 0.109 * | 0.024 | 0.070 | 0.025 |
| Hemojuvelin BMP co-receptor (RGM-C) | Other | DASH | 0.090 | 0.024 | 0.105 * | 0.025 | 0.088 | 0.026 |
| Heterogeneous nuclear ribonucleoprotein A/B (hnRNP A/B) | Enzyme | DASH | −0.054 | 0.023 | −0.099 * | 0.024 | −0.042 | 0.025 |
| Kynureninase (KYNU) | Enzyme | AHEI, MDS, DASH | −0.127 * | 0.024 | −0.153 * | 0.024 | −0.112 * | 0.026 |
| Leucine rich repeats and immunoglobulin like domains 3 (LRIG3) | Other | AHEI | 0.114 * | 0.024 | 0.087 | 0.025 | 0.069 | 0.026 |
| Macrophage migration inhibitory factor (MIF) | MDS, DASH | −0.098 | 0.024 | −0.119 * | 0.024 | −0.106 * | 0.026 | |
| Matrix metallopeptidase 2 (MMP-2) | Peptidase | DASH | 0.076 | 0.024 | 0.117 * | 0.024 | 0.074 | 0.025 |
| MET proto-oncogene, receptor tyrosine kinase (MET) | Kinase | AHEI | 0.102 * | 0.023 | 0.096 | 0.024 | 0.050 | 0.025 |
| Notch 1 | Transcription regulator | AHEI, DASH | 0.098 * | 0.023 | 0.113 * | 0.023 | 0.069 | 0.024 |
| Peroxiredoxin 1 | Enzyme | DASH | −0.068 | 0.024 | −0.113 * | 0.025 | −0.077 | 0.026 |
| Plasminogen activator, tissue type (tPA) | Peptidase | DASH | −0.048 | 0.022 | −0.101 * | 0.022 | −0.068 | 0.023 |
| Serpin family C member 1 (Antithrombin III) | Enzyme | AHEI | 0.093 * | 0.021 | 0.063 | 0.022 | 0.053 | 0.023 |
| Stanniocalcin 1 | Kinase | AHEI, MDS, DASH | −0.108 * | 0.024 | −0.103 * | 0.024 | −0.111 * | 0.025 |
| Thrombospondin 2 (TSP2) | Other | AHEI | −0.112 * | 0.024 | −0.087 | 0.025 | −0.077 | 0.026 |
| Transferrin | Transporter | AHEI | 0.127 * | 0.022 | 0.083 | 0.023 | 0.068 | 0.024 |
| WAP, follistatin/kazal, immunoglobulin, kunitz and netrin domain containing 2 (WFKN2) | Other | AHEI, DASH | 0.095 * | 0.023 | 0.097 * | 0.023 | 0.099 | 0.024 |
| Prostatic binding protein | DASH | −0.046 | 0.024 | −0.101 * | 0.024 | −0.042 | 0.026 | |
| Name | Type\Class | Diet 2 | AHEI | DASH | MDS | |||
|---|---|---|---|---|---|---|---|---|
| Β 3 | SE | β | SE | β | SE | |||
| Aconitate | Aliphatic acyclic compound | AHEI, DASH | −0.025 * | 0.006 | −0.025 * | 0.006 | −0.019 | 0.007 |
| Cholesterol ester (C22:6) | Lipid | AHEI, MDS | 0.059 * | 0.015 | 0.048 | 0.016 | 0.080 * | 0.017 |
| Cis/trans-hydroxyproline | L-alpha-amino acid | MDS, DASH | −0.033 | 0.012 | −0.070 * | 0.012 | −0.054 * | 0.013 |
| Cotinine | MDS, DASH | −2.094 | 0.723 | −3.075 * | 0.734 | −3.097 * | 0.772 | |
| Hippurate | Aromatic homomonocyclic compounds | AHEI, MDS, DASH | 0.364 * | 0.079 | 0.430 * | 0.080 | 0.354 * | 0.085 |
| Isocitrate | Aliphatic acyclic compounds | AHEI, MDS, DASH | −0.041 * | 0.008 | −0.049 * | 0.008 | −0.041 * | 0.008 |
| Lysophosphatidylcholine (C20:5) | Lipid | AHEI, DASH | 0.079 * | 0.016 | 0.069 * | 0.017 | 0.062 | 0.018 |
| Lysophosphatidylcholine (C22:6) | Lipid | AHEI, MDS, DASH | 0.073 * | 0.010 | 0.052 * | 0.011 | 0.077 * | 0.011 |
| Lysophosphatidylethanolamine (C20:4) | Lipid | MDS | −0.022 | 0.009 | −0.028 | 0.009 | −0.045 * | 0.010 |
| Ornithine | L-alpha-amino acid | DASH | −0.028 | 0.011 | −0.057 * | 0.011 | −0.039 | 0.012 |
| Oxalate | Aliphatic acyclic compounds | MDS, DASH | 0.064 | 0.018 | 0.107 * | 0.018 | 0.086 * | 0.019 |
| Pantothenate | Water-soluble vitamins | DASH | 0.049 | 0.022 | 0.126 * | 0.022 | 0.083 | 0.023 |
| Phosphatidylcholine (C38:6) | Lipid | AHEI, MDS, DASH | 0.039 * | 0.005 | 0.035 * | 0.005 | 0.046 * | 0.005 |
| Phosphatidylcholine (40:6) | Lipid | AHEI, MDS, DASH | 0.041 * | 0.006 | 0.038 * | 0.006 | 0.054 * | 0.006 |
| Serine | Amino acid | DASH | −0.006 | 0.007 | −0.030 * | 0.007 | −0.018 | 0.008 |
| Sphingomyelin (C14:0) | Lipid | MDS | −0.012 | 0.006 | −0.001 | 0.007 | −0.043 * | 0.007 |
| Sphingomyelin (C18:0) | Lipid | AHEI, MDS, DASH | −0.026 * | 0.006 | −0.028 * | 0.006 | −0.032 * | 0.006 |
| Sphingomyelin (C18:1) | Lipid | AHEI, MDS, DASH | −0.044 * | 0.007 | −0.044 * | 0.007 | −0.036 * | 0.007 |
| Thiamine | Water-soluble vitamins | DASH | 0.067 | 0.037 | 0.150 * | 0.037 | 0.101 | 0.039 |
| Triacylglycerol (C54:7) | Lipid | MDS | 0.036 | 0.012 | 0.037 | 0.012 | 0.050 * | 0.013 |
| Triacylglycerol (C56:7) | Lipid | AHEI, MDS, DASH | 0.037 * | 0.007 | 0.031 * | 0.007 | 0.041 * | 0.008 |
| Triacylglycerol (C56:8) | Lipid | AHEI, MDS, DASH | 0.043 * | 0.009 | 0.040 * | 0.009 | 0.054 * | 0.009 |
| Triacylglycerol (C58:10) | Lipid | AHEI, MDS, DASH | 0.045 * | 0.010 | 0.038 * | 0.010 | 0.048 * | 0.011 |
| Triacylglycerol (C58:8) | Lipid | AHEI, MDS, DASH | 0.042 * | 0.008 | 0.037 * | 0.008 | 0.047 * | 0.008 |
| Triacylglycerol (C58:9) | Lipid | AHEI, MDS, DASH | 0.050 * | 0.009 | 0.045 * | 0.009 | 0.057 * | 0.009 |
| Uridine | Ribonucleosides | AHEI, DASH | 0.028 * | 0.005 | 0.034 * | 0.005 | 0.019 | 0.005 |
| Pathway | Enrichment Ratio | P-Value 2 | FDR Q-Value 3 | Matched Molecules | |
|---|---|---|---|---|---|
| hsa04610 | Complement and coagulation cascades | 12.51 | 3.1 × 108 | 1.0 × 105 | PLAT, SERPINC1, C3, SERPINE1, SERPINA5, CFH, C1S, C3AR1, CPB2 |
| hsa05150 | Staphylococcus aureus infection | 9.81 | 1.4 × 104 | 1.7 × 102 | C3, CFH, C1S, SELP |
| hsa01522 | Endocrine resistance | 6.72 | 2.5 × 104 | 1.7 × 102 | EGFR, MMP2, BCL2, NOTCH1, MED1, JAG1 |
| hsa04066 | HIF-1 signaling pathway | 6.59 | 2.8 × 104 | 1.7 × 102 | EGFR, GAPDH, SERPINE1, TEK, TF, BCL2 |
| hsa05230 | Central carbon metabolism in cancer | 8.45 | 2.9 × 104 | 1.7 × 102 | EGFR, FGFR1, KIT, MET, NTRK3 |
| hsa04151 | PI3K-Akt signaling pathway | 3.41 | 3.1 × 104 | 1.7 × 102 | CDC37, EGFR, FGFR1, HSP90AB1, MET, TEK, THBS2, BCL2, KIT, YWHAB |
| hsa05144 | Malaria | 8.97 | 9.8 × 104 | 4.6 × 102 | MET, SELE, SELP, THBS2 |
| hsa05418 | Fluid shear stress and atherosclerosis | 4.74 | 1.6 × 103 | 6.2 × 102 | HSP90AB1, MMP2, PLAT, BCL2, SELE, BMPR1A |
| hsa05215 | Prostate cancer | 5.66 | 1.8 × 103 | 6.2 × 102 | EGFR, PLAT, HSP90AB1, FGFR1, BCL2 |
| hsa04514 | Cell adhesion molecules (CAMs) | 4.58 | 1.9 × 103 | 6.2 × 102 | SELP, SELE, CNTN1, NEGR1, PDCD1LG2, NRXN3 |
| Protein 1 | Variant 2 | Nearest Gene | GWAS Associated Traits 3 | GWAS Study Accession 4 |
|---|---|---|---|---|
| AK1A1 | rs72688441 | NASP | Blood protein concentrations | GCST006585, GCST005806 |
| AK1A1 | 1:46767127:TCTC_ | LRRC41 | ||
| AK1A1 | rs72684498 | ZSWIM5 | ||
| AK1A1 | rs72676591 | EIF2B3 | ||
| Antithrombin III | rs334516 | TNS3 | ||
| bFGF-R | 8:38324424:ACC_A | FGFR1 | ||
| C1s | rs6695321 | CFH | Blood protein concentrations | GCST004365, GCST005806 |
| C1s | rs150845796 | CFHR2 | ||
| C3a | rs11583804 | ACTRT2 | ||
| Cadherin-5 | rs8176672 | ABO | Blood protein concentrations, optic cup area | GCST005806, GCST004137 |
| Cadherin-5 | rs11534419 | TMEM8C | ||
| Carbonic anhydrase 6 | rs11576766 | CA6 | ||
| Cathepsin D | rs1558500 | |||
| Cathepsin S | rs72702561 | HORMAD1 | Blood protein concentrations | GCST006585 |
| Cathepsin S | rs9661107 | PRUNE1 | ||
| Cathepsin S | rs141935877 | TARS2 | ||
| Contactin-1 | rs11640313 | MRPL28 | ||
| ERBB1 | rs12112554 | |||
| Factor H | rs10737680 | CFH | Macular degeneration, lung function, blood protein concentrations | GCST004737, GCST003265, GCST004365 |
| Factor H | rs12134610 | KCNT2 | ||
| Factor H | rs150845796 | CFHR2 | ||
| Factor H | rs1329428 | Macular degeneration, central serous retinopathy, lung function | GCST000806, GCST006416, GCST003265 | |
| Gal 3 | rs11733361 | SLC25A31 | ||
| iC3b | rs11583804 | ACTRT2 | ||
| IGFBP-1 | rs139504202 | |||
| KYNU | rs35647509 | KYNU | ||
| KYNU | rs4733300 | NRG1 | ||
| LSAMP | rs117955663 | OR56B2P | ||
| MET | rs635634 | ABO | Blood protein concentrations, LDL-C concentrations, TC concentrations, ischemic stroke, immune cell count, T2D | GCST000759, GCST000760, GCST006910, GCST004613, GCST004773, GCST005806 |
| Notch 1 | rs138015312 | UGT2B15 | ||
| PARC | rs1102934 | CCL18 | Blood protein concentrations | GCST006585 |
| RGM-C | rs2381409 | FAM221B | ||
| RGM-C | 7:57407593:TTCC_ | |||
| sE-Selectin | rs2519093 | ABO | Blood protein concentrations, LDL-C concentrations, venous thromboembolism, TC concentrations, hematocrit, hemoglobin, immune cell count, CAD | GCST005806, GCST006612, GCST004256, GCST007143, GCST005994, GCST005995, GCST007070, GCST005195 |
| sE-Selectin | rs9722289 | TMEM8C | ||
| TIG2 | rs10282458 | LOC107986858 | ||
| Transferrin | rs71544591 | PTPRN2 | ||
| TSP2 | rs73043837 | THBS2 | ||
| WFKN2 | rs7207028 | WFIKKN2 |
| Metabolite 1 | Variant 2 | Nearest Gene | GWAS Associated Traits 3 | GWAS Study Accession4 |
|---|---|---|---|---|
| Cholesterol ester (C20:5) | rs174548 | FADS1-2 | Blood metabolite concentrations, metabolite ratios in CKD, TG concentrations, HDL-C concentrations, plasma N6 concentrations, metabolite ratios | GCST000274, GCST005650, GCST000809, GCST000805, GCST003237, GCST002442 |
| Indoxylsulfate | rs875480 | CSNK1G3 | ||
| Lysophosphatidylcholine (C20:5) | rs174548 | FADS1-2 | Blood metabolite concentrations, metabolite ratios in CKD, TG concentrations, HDL-C concentrations, plasma N6 concentrations, metabolite ratios | GCST000274, GCST005650, GCST000809, GCST000805, GCST003237, GCST002442 |
| Lysophosphatidylcholine (C22:6) | rs174550 | FADS1-2 | Blood metabolite concentrations, TG concentrations, FBG, HDL-C concentrations, LDL-C concentrations, plasma N6 concentrations, plasma N3 concentrations, HOMA-B | GCST002443, GCST004237. GCST005186, GCST004232, GCST007141, GCST002446, GCST001178, GCST005180, |
| Lysophosphatidylethanolamine (C18:2) | rs4246215 | FADS2 | Plasma N3 concentrations, platelet count, IBD, RBC fatty acid concentrations, Crohn’s disease, CRC | GCST001180, GCST001337, GCST001725, GCST002712, GCST004132, GCST007992 |
| Lysophosphatidylethanolamine (C20:4) | rs174548 | FADS1-2 | Blood metabolite concentrations, metabolite ratios in CKD, TG concentrations, HDL-C concentrations, plasma N6 concentrations, metabolite ratios | GCST000274, GCST005650, GCST000809, GCST000805, GCST003237, GCST002442 |
| Lysophosphatidylethanolamine (C20:4) | rs4149056 | SLCO1B1 | Blood metabolite concentrations, TG concentrations, bilirubin, response to statin, ER+ breast cancer, thyroxine concentrations, hemoglobin, metabolite ratios | GCST002442, GCST007133, GCST000386, GCST000213, GCST004359, GCST006896, GCST007068, GCST002442 |
| Phosphatidylcholine (C36:2) | rs174541 | FADS2 | Blood metabolite concentrations, metabolite ratios in CKD, trans fatty acid concentrations, RBC fatty acids, | GCST001852, GCST005650, GCST002721, GCST002712 |
| Phosphatidylcholine (C40:6) | rs174535 | FADS1-2 | Blood metabolite concentrations, plasma N3 concentrations, RBC fatty acids, trans fatty acid concentrations, inflammatory disease, asthma, glycemic traits, respiratory disease | GCST002443, GCST001178, GCST002712, GCST002721, GCST005537, GCST007799, GCST008674, GCST007076 |
| Proline | rs2078743 | PRODH | ||
| Serine | rs477992 | PHGDH | Blood metabolite/amino acid concentrations, TC concentrations | GCST002966, GCST006614 |
| Sphingomyelin (C14:0) | rs11158519 | SYNE2 | Sphingomyelin concentrations | GCST008933 |
| Triacylglycerol (C54:2) | rs964184 | APOA1/C3/A4/A5 | Blood metabolite concentrations, TG concentrations, lipoprotein concentrations, CHD, stroke, MetS, phospholipid fatty acids, fat soluble vitamin concentrations, CAD, age related CVDs | GCST001639, GCST004550, GCST004759, GCST000998, GCST002290, GCST001436, GCST001414, GCST001142, GCST004787, GCST004045, |
| Triacylglycerol (C54:4) | rs174550 | FADS1-2 | TG concentrations, FBG, HDL-C concentrations, LDL-C concentrations, plasma N6 concentrations, HOMA-B, plasma N3 concentrations, RBC fatty acids, trans fatty acid concentrations | GCST004237, GCST008032, GCST004232, GCST007141, GCST002448, GCST005180, GCST001178, GCST002712, GCST002721 |
| Triacylglycerol (C54:5) | rs964184 | APOA1/C3/A4/A5 | Blood metabolite concentrations, TG concentrations, lipoprotein concentrations, CHD, stroke,, MetS, phospholipid fatty acids, fat soluble vitamin concentrations, CAD, age related CVDs | GCST001639, GCST004550, GCST004759, GCST000998, GCST002290, GCST001436, GCST001414, GCST001142, GCST004787, GCST004045, |
| Triacylglycerol (C56:7) | rs6593086 | |||
| Triacylglycerol (C58:10) | rs174548 | FADS1-2 | Blood metabolite concentrations, metabolite ratios in CKD, TG concentrations, HDL-C concentrations, plasma N6 concentrations, metabolite ratios | GCST000274, GCST005650, GCST000809, GCST000805, GCST003237, GCST002442 |
| Triacylglycerol (C58:8) | rs6593086 | |||
| Triacylglycerol (C58:9) | rs6593086 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walker, M.E.; Song, R.J.; Xu, X.; Gerszten, R.E.; Ngo, D.; Clish, C.B.; Corlin, L.; Ma, J.; Xanthakis, V.; Jacques, P.F.; et al. Proteomic and Metabolomic Correlates of Healthy Dietary Patterns: The Framingham Heart Study. Nutrients 2020, 12, 1476. https://doi.org/10.3390/nu12051476
Walker ME, Song RJ, Xu X, Gerszten RE, Ngo D, Clish CB, Corlin L, Ma J, Xanthakis V, Jacques PF, et al. Proteomic and Metabolomic Correlates of Healthy Dietary Patterns: The Framingham Heart Study. Nutrients. 2020; 12(5):1476. https://doi.org/10.3390/nu12051476
Chicago/Turabian StyleWalker, Maura E., Rebecca J. Song, Xiang Xu, Robert E. Gerszten, Debby Ngo, Clary B. Clish, Laura Corlin, Jiantao Ma, Vanessa Xanthakis, Paul F. Jacques, and et al. 2020. "Proteomic and Metabolomic Correlates of Healthy Dietary Patterns: The Framingham Heart Study" Nutrients 12, no. 5: 1476. https://doi.org/10.3390/nu12051476