Inositol and Non-Alcoholic Fatty Liver Disease: A Systematic Review on Deficiencies and Supplementation

 , , , , , and
, , , , , and
Abstract
:1. Introduction
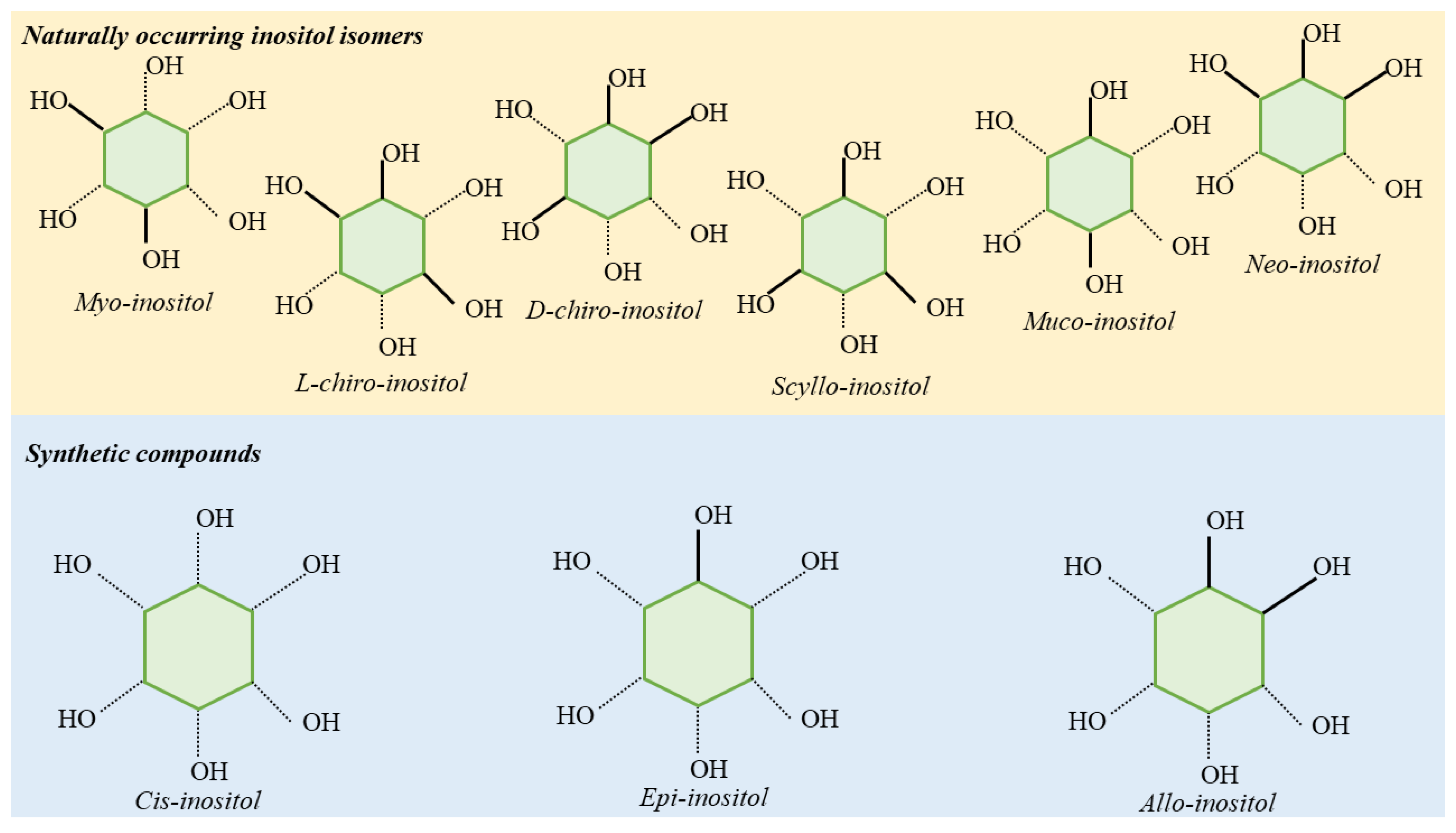
2. Biological Role of Inositol and Its Derivates
3. Inositol Pharmacology
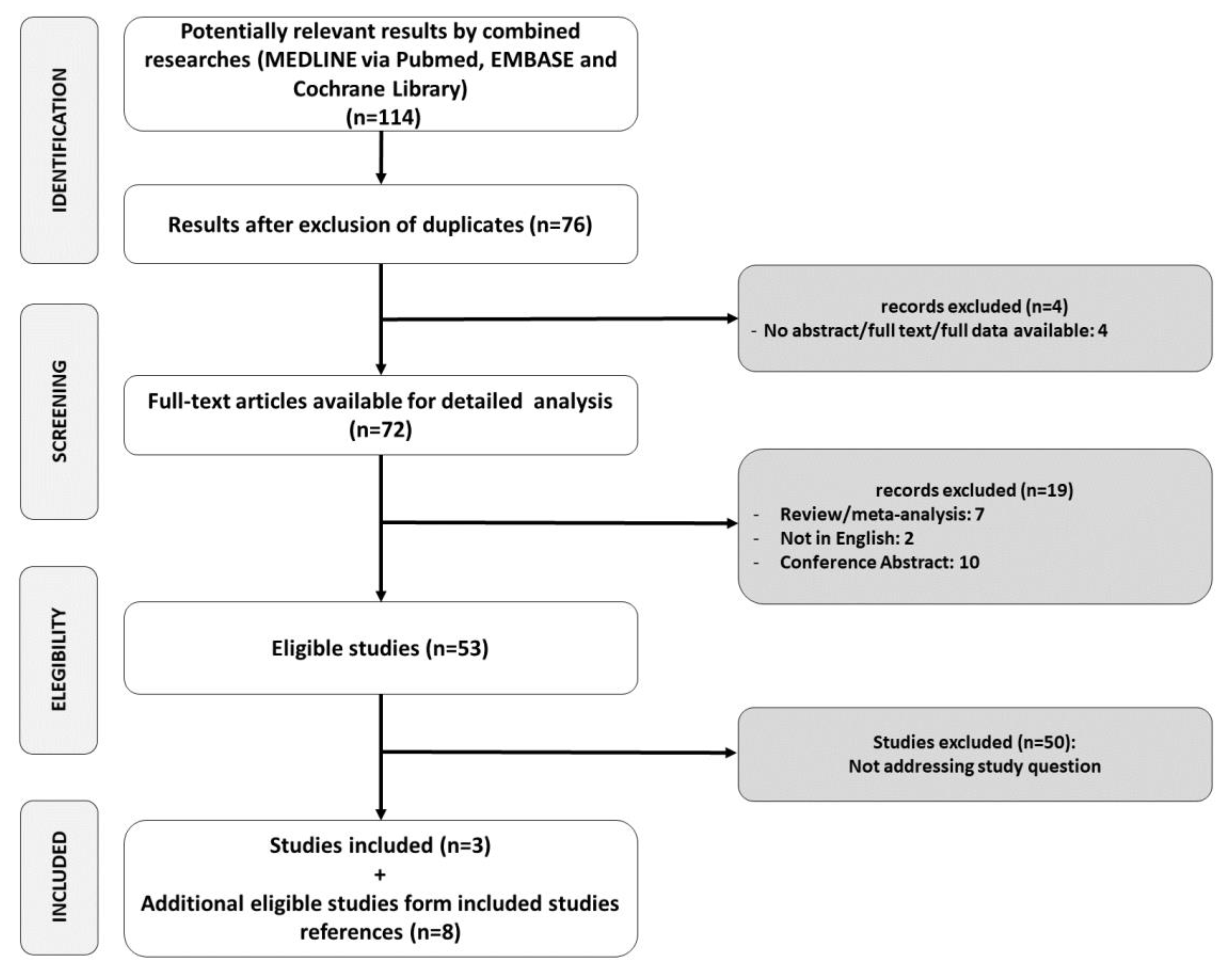
4. Systematic Review Methods
4.1. Eligibility Criteria
4.2. Information Sources and Search Strategy
4.3. Study Selection
4.4. Data Collection Process and Data Items
4.5. Ethical Review
5. Results
6. Inositol Deficiency in NAFLD
7. Inositol Supplementation in NAFLD
8. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of NAFLD and NASH: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef]
- Charlton, M.R.; Burns, J.M.; Pedersen, R.A.; Watt, K.D.; Heimbach, J.K.; Dierkhising, R.A. Frequency and outcomes of liver transplantation for nonalcoholic steatohepatitis in the United States. Gastroenterology 2011, 141, 1249–1253. [Google Scholar] [CrossRef]
- Baratta, F.; Pastori, D.; Angelico, F.; Balla, A.; Paganini, A.M.; Cocomello, N.; Ferro, D.; Violi, F.; Sanyal, A.J.; Del Ben, M. Nonalcoholic Fatty Liver Disease and Fibrosis Associated With Increased Risk of Cardiovascular Events in a Prospective Study. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2020, 18, 2324–2331.e4. [Google Scholar] [CrossRef]
- Anstee, Q.M.; Mantovani, A.; Tilg, H.; Targher, G. Risk of cardiomyopathy and cardiac arrhythmias in patients with nonalcoholic fatty liver disease. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 425–439. [Google Scholar] [CrossRef]
- Hydes, T.; Buchanan, R.; Kennedy, O.J.; Fraser, S.; Parkes, J.; Roderick, P. Systematic review of the impact of non-alcoholic fatty liver disease on mortality and adverse clinical outcomes for individuals with chronic kidney disease. BMJ Open 2020, 10, e040970. [Google Scholar] [CrossRef]
- Kumar, J.; Memon, R.S.; Shahid, I.; Rizwan, T.; Zaman, M.; Menezes, R.G.; Kumar, S.; Siddiqi, T.J.; Usman, M.S. Antidiabetic drugs and non-alcoholic fatty liver disease: A systematic review, meta-analysis and evidence map. Dig. Liver Dis. Off. J. Ital. Soc. Gastroenterol. Ital. Assoc. Study Liver 2020. [Google Scholar] [CrossRef]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef]
- Balla, T.; Bondeva, T.; Várnai, P. How accurately can we image inositol lipids in living cells? Trends Pharmacol. Sci. 2000, 21, 238–241. [Google Scholar] [CrossRef]
- Wilcox, R.A. Measurement of calcium fluxes in permeabilized cells using a 45Ca2+ uptake and release assay. Methods Mol. Biol. 2005, 312, 205–212. [Google Scholar]
- Pani, A.; Gironi, I.; Di Vieste, G.; Mion, E.; Bertuzzi, F.; Pintaudi, B. From Prediabetes to Type 2 Diabetes Mellitus in Women with Polycystic Ovary Syndrome: Lifestyle and Pharmacological Management. Int. J. Endocrinol. 2020, 2020, 6276187. [Google Scholar] [CrossRef]
- Thomas, R.M.; Nechamen, C.A.; Mazurkiewicz, J.E.; Ulloa-Aguirre, A.; Dias, J.A. The adapter protein APPL1 links FSH receptor to inositol 1,4,5-trisphosphate production and is implicated in intracellular Ca(2+) mobilization. Endocrinology 2011, 152, 1691–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pace, C.; Tumino, D.; Russo, M.; Le Moli, R.; Naselli, A.; Borzì, G.; Malandrino, P.; Frasca, F. Role of selenium and myo-inositol supplementation on autoimmune thyroiditis progression. Endocr. J. 2020. [Google Scholar] [CrossRef]
- Chiu, T.T.Y.; Rogers, M.S.; Briton-Jones, C.; Haines, C. Effects of myo-inositol on the in-vitro maturation and subsequent development of mouse oocytes. Hum. Reprod. 2003, 18, 408–416. [Google Scholar] [CrossRef] [Green Version]
- Croze, M.L.; Soulage, C.O. Potential role and therapeutic interests of myo-inositol in metabolic diseases. Biochimie 2013, 95, 1811–1827. [Google Scholar] [CrossRef]
- Caputo, M.; Bona, E.; Leone, I.; Samà, M.T.; Nuzzo, A.; Ferrero, A.; Aimaretti, G.; Marzullo, P.; Prodam, F. Inositols and metabolic disorders: From farm to bedside. J. Tradit. Complement. Med. 2020, 10, 252–259. [Google Scholar] [CrossRef]
- Hipps, P.P.; Sehgal, R.K.; Holland, W.H.; Sherman, W.R. Identification and partial characterization of inositol: NAD+ epimerase and inosose: NAD(P)H reductase from the fat body of the American cockroach, Periplaneta americana L. Biochemistry 1973, 12, 4507–4512. [Google Scholar] [CrossRef]
- Streb, H.; Irvine, R.F.; Berridge, M.J.; Schulz, I. Release of Ca2+ from a nonmitochondrial intracellular store in pancreatic acinar cells by inositol-1,4,5-trisphosphate. Nature 1983, 306, 67–69. [Google Scholar] [CrossRef]
- Larner, J.; Craig, J.W. Urinary myo-inositol-to-chiro-inositol ratios and insulin resistance. Diabetes Care 1996, 19, 76–78. [Google Scholar] [CrossRef]
- Santamaria, A.; Giordano, D.; Corrado, F.; Pintaudi, B.; Interdonato, M.L.; Vieste, G.D.; Di Benedetto, A.; D’Anna, R. One-year effects of myo-inositol supplementation in postmenopausal women with metabolic syndrome. Climacteric 2012, 15, 490–495. [Google Scholar] [CrossRef]
- Pintaudi, B.; Di Vieste, G.; Bonomo, M. The Effectiveness of Myo-Inositol and D-Chiro Inositol Treatment in Type 2 Diabetes. Int. J. Endocrinol. 2016, 2016, 9132052. [Google Scholar] [CrossRef] [Green Version]
- Maeba, R.; Hara, H.; Ishikawa, H.; Hayashi, S.; Yoshimura, N.; Kusano, J.; Takeoka, Y.; Yasuda, D.; Okazaki, T.; Kinoshita, M.; et al. Myo-inositol treatment increases serum plasmalogens and decreases small dense LDL, particularly in hyperlipidemic subjects with metabolic syndrome. J. Nutr. Sci. Vitaminol. 2008, 54, 196–202. [Google Scholar] [CrossRef] [Green Version]
- Nordio, M.; Proietti, E. The combined therapy with myo-inositol and D-chiro-inositol reduces the risk of metabolic disease in PCOS overweight patients compared to myo-inositol supplementation alone. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 575–581. [Google Scholar]
- Capasso, I.; Esposito, E.; Maurea, N.; Montella, M.; Crispo, A.; De Laurentiis, M.; D’Aiuto, M.; Frasci, G.; Botti, G.; Grimaldi, M.; et al. Combination of inositol and alpha lipoic acid in metabolic syndrome-affected women: A randomized placebo-controlled trial. Trials 2013, 14, 273. [Google Scholar] [CrossRef] [Green Version]
- Fruzzetti, F.; Perini, D.; Russo, M.; Bucci, F.; Gadducci, A. Comparison of two insulin sensitizers, metformin and myo-inositol, in women with polycystic ovary syndrome (PCOS). Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2017, 33, 39–42. [Google Scholar] [CrossRef]
- Grases, F.; Simonet, B.M.; Vucenik, I.; Prieto, R.M.; Costa-Bauzá, A.; March, J.G.; Shamsuddin, A.M. Absorption and excretion of orally administered inositol hexaphosphate (IP(6) or phytate) in humans. Biofactors 2001, 15, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Carlomagno, G.; De Grazia, S.; Unfer, V.; Manna, F. Myo-inositol in a new pharmaceutical form: A step forward to a broader clinical use. Expert Opin. Drug Deliv. 2012, 9, 267–271. [Google Scholar] [CrossRef]
- Holub, B.J. 1982 Borden Award lecture. Nutritional, biochemical, and clinical aspects of inositol and phosphatidylinositol metabolism. Can. J. Physiol. Pharmacol. 1984, 62, 1–8. [Google Scholar] [CrossRef]
- Papaleo, E.; Unfer, V.; Baillargeon, J.-P.; Fusi, F.; Occhi, F.; De Santis, L. Myo-inositol may improve oocyte quality in intracytoplasmic sperm injection cycles. A prospective, controlled, randomized trial. Fertil. Steril. 2009, 91, 1750–1754. [Google Scholar] [CrossRef]
- Barak, Y.; Levine, J.; Glasman, A.; Elizur, A.; Belmaker, R.H. Inositol treatment of Alzheimer’s disease: A double blind, cross-over placebo controlled trial. Prog. Neuropsychopharmacol. Biol. Psychiatry 1996, 20, 729–735. [Google Scholar] [CrossRef]
- Vitagliano, A.; Saccone, G.; Cosmi, E.; Visentin, S.; Dessole, F.; Ambrosini, G.; Berghella, V. Inositol for the prevention of gestational diabetes: A systematic review and meta-analysis of randomized controlled trials. Arch. Gynecol. Obstet. 2019, 299, 55–68. [Google Scholar] [CrossRef]
- Carlomagno, G.; Unfer, V. Inositol safety: Clinical evidences. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 931–936. [Google Scholar]
- Özturan, A.; Arslan, S.; Kocaadam, B.; Elibol, E.; İmamoğlu, İ.; Karadağ, M.G. Effect of inositol and its derivatives on diabetes: A systematic review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1124–1136. [Google Scholar] [CrossRef] [PubMed]
- Pintaudi, B.; Di Vieste, G.; Corrado, F.; Lucisano, G.; Giunta, L.; D’Anna, R.; Di Benedetto, A. Effects of myo-inositol on glucose variability in women with gestational diabetes. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 6567–6572. [Google Scholar]
- Fraticelli, F.; Celentano, C.; Zecca, I.A.; Di Vieste, G.; Pintaudi, B.; Liberati, M.; Franzago, M.; Di Nicola, M.; Vitacolonna, E. Effect of inositol stereoisomers at different dosages in gestational diabetes: An open-label, parallel, randomized controlled trial. Acta Diabetol. 2018, 55, 805–812. [Google Scholar] [CrossRef]
- Laganà, A.S.; Garzon, S.; Casarin, J.; Franchi, M.; Ghezzi, F. Inositol in Polycystic Ovary Syndrome: Restoring Fertility through a Pathophysiology-Based Approach. Trends Endocrinol. Metab. 2018, 29, 768–780. [Google Scholar] [CrossRef]
- Hayashi, E.; Maeda, T.; Tomita, T. The effect of myo-inositol deficiency on lipid metabolism in rats. I. The alteration of lipid metabolism in myo-inositol deficient rats. Biochim. Biophys. Acta 1974, 360, 134–145. [Google Scholar] [CrossRef]
- Hayashi, E.; Maeda, T.; Tomita, T. The effect of myo-inositol deficiency on lipid metabolism in rats. II. The mechanism of triacylglycerol accumulation in the liver of myo-inositol-deficient rats. Biochim. Biophys. Acta 1974, 360, 146–155. [Google Scholar] [CrossRef]
- Thakur, P.C.; Stuckenholz, C.; Rivera, M.R.; Davison, J.M.; Yao, J.K.; Amsterdam, A.; Sadler, K.C.; Bahary, N. Lack of de novo phosphatidylinositol synthesis leads to endoplasmic reticulum stress and hepatic steatosis in cdipt-deficient zebrafish. Hepatology 2011, 54, 452–462. [Google Scholar] [CrossRef] [Green Version]
- Katayama, T. Effect of dietary addition of myo-inositol on lipid metabolism in rats fed sucrose or corn starch. Nutr. Res. 1994, 14, 699–706. [Google Scholar] [CrossRef]
- Katayama, T. Effects of dietary myo-inositol or phytic acid on hepatic concentrations of lipids and hepatic activities of lipogenic enzymes in rats fed on corn starch or sucrose. Nutr. Res. 1997, 17, 721–728. [Google Scholar] [CrossRef]
- Geethan, P.K.M.A.; Prince, P.S.M. Antihyperlipidemic effect of D-pinitol on streptozotocin-induced diabetic Wistar rats. J. Biochem. Mol. Toxicol. 2008, 22, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Park, C.-M.; Cho, C.-W.; Song, Y.-S. Protective Effect of Pinitol against D-Galactosamine-Induced Hepatotoxicity in Rats Fed on a High-Fat Diet. Biosci. Biotechnol. Biochem. 2008, 72, 1657–1666. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.-S.; Lee, M.-K.; Jung, U.J.; Kim, H.-J.; Do, G.-M.; Park, Y.B.; Jeon, S.-M. Metabolic response of soy pinitol on lipid-lowering, antioxidant and hepatoprotective action in hamsters fed-high fat and high cholesterol diet. Mol. Nutr. Food Res. 2009, 53, 751–759. [Google Scholar] [CrossRef]
- Sivakumar, S.; Palsamy, P.; Subramanian, S.P. Attenuation of oxidative stress and alteration of hepatic tissue ultrastructure by D-pinitol in streptozotocin-induced diabetic rats. Free Radic. Res. 2010, 44, 668–678. [Google Scholar] [CrossRef]
- Shimada, M.; Ichigo, Y.; Shirouchi, B.; Takashima, S.; Inagaki, M.; Nakagawa, T.; Hayakawa, T. Treatment with myo-inositol attenuates binding of the carbohydrate-responsive element-binding protein to the ChREBP-β and FASN genes in rat nonalcoholic fatty liver induced by high-fructose diet. Nutr. Res. 2019, 64, 49–55. [Google Scholar] [CrossRef]
- Lee, E.; Lim, Y.; Kwon, S.W.; Kwon, O. Pinitol consumption improves liver health status by reducing oxidative stress and fatty acid accumulation in subjects with non-alcoholic fatty liver disease: A randomized, double-blind, placebo-controlled trial. J. Nutr. Biochem. 2019, 68, 33–41. [Google Scholar] [CrossRef]
- Del Ben, M.; Polimeni, L.; Baratta, F.; Pastori, D.; Angelico, F. The role of nutraceuticals for the treatment of non-alcoholic fatty liver disease. Br. J. Clin. Pharmacol. 2017, 83, 88–95. [Google Scholar] [CrossRef]
- Lombardi, R.; Onali, S.; Thorburn, D.; Davidson, B.R.; Gurusamy, K.S.; Tsochatzis, E. Pharmacological interventions for non-alcohol related fatty liver disease (NAFLD): An attempted network meta-analysis. Cochrane Database Syst. Rev. 2017, 3, CD011640. [Google Scholar]
- Pastori, D.; Polimeni, L.; Baratta, F.; Pani, A.; Del Ben, M.; Angelico, F. The efficacy and safety of statins for the treatment of non-alcoholic fatty liver disease. Dig. Liver Dis. Off. J. Ital. Soc. Gastroenterol. Ital. Assoc. Study Liver 2015, 47, 4–11. [Google Scholar] [CrossRef] [Green Version]
- Del Ben, M.; Polimeni, L.; Baratta, F.; Pastori, D.; Loffredo, L.; Angelico, F. Modern approach to the clinical management of non-alcoholic fatty liver disease. World J. Gastroenterol. 2014, 20, 8341–8350. [Google Scholar]
- Chu, S.W.; Geyer, R.P. myo-Inositol action on gerbil intestine. Association of phosphatidylinositol metabolism with lipid clearance. Biochim. Biophys. Acta 1982, 710, 63–70. [Google Scholar] [CrossRef]
- McCrea, H.J.; De Camilli, P. Mutations in phosphoinositide metabolizing enzymes and human disease. Physiology 2009, 24, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Muscogiuri, G.; Palomba, S.; Laganà, A.S.; Orio, F. Inositols in the Treatment of Insulin-Mediated Diseases. Int. J. Endocrinol. 2016, 2016, 3058393. [Google Scholar] [CrossRef] [Green Version]
- Ozcan, U.; Cao, Q.; Yilmaz, E.; Lee, A.-H.; Iwakoshi, N.N.; Ozdelen, E.; Tuncman, G.; Görgün, C.; Glimcher, L.H.; Hotamisligil, G.S. Endoplasmic reticulum stress links obesity, insulin action, and type 2 diabetes. Science 2004, 306, 457–461. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Tan, M.; Hu, Y.; Wang, J.-L.; Scheuner, D.; Kaufman, R.J. Ultraviolet light activates NFkappaB through translational inhibition of IkappaBalpha synthesis. J. Biol. Chem. 2004, 279, 34898–34902. [Google Scholar] [CrossRef] [Green Version]
- Malhi, H.; Kaufman, R.J. Endoplasmic reticulum stress in liver disease. J. Hepatol. 2011, 54, 795–809. [Google Scholar] [CrossRef] [Green Version]
- Sanyal, A.J. Past, present and future perspectives in nonalcoholic fatty liver disease. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Noventa, M.; Vitagliano, A.; Quaranta, M.; Borgato, S.; Abdulrahim, B.; Gizzo, S. Preventive and Therapeutic Role of Dietary Inositol Supplementation in Periconceptional Period and During Pregnancy: A Summary of Evidences and Future Applications. Reprod. Sci. 2016, 23, 278–288. [Google Scholar] [CrossRef]
- Bizzarri, M.; Cucina, A.; Dinicola, S.; Harrath, A.H.; Alwasel, S.H.; Unfer, V.; Bevilacqua, A. Does myo-inositol effect on PCOS follicles involve cytoskeleton regulation? Med. Hypotheses 2016, 91, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Michell, R.H. Do inositol supplements enhance phosphatidylinositol supply and thus support endoplasmic reticulum function? Br. J. Nutr. 2018, 120, 301–316. [Google Scholar] [CrossRef] [Green Version]
- Beyoğlu, D.; Imbeaud, S.; Maurhofer, O.; Bioulac-Sage, P.; Zucman-Rossi, J.; Dufour, J.-F.; Idle, J.R. Tissue metabolomics of hepatocellular carcinoma: Tumor energy metabolism and the role of transcriptomic classification. Hepatology 2013, 58, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Ignat, M.; Akladios, C.Y.; Lindner, V.; Khetchoumian, K.; Teletin, M.; Muttter, D.; Aprahamian, P.M.; Marescaux, J. Development of a methodology for in vivo follow-up of hepatocellular carcinoma in hepatocyte specific Trim24-null mice treated with myo-inositol trispyrophosphate. J. Exp. Clin. Cancer Res. 2016, 35, 155. [Google Scholar] [CrossRef] [Green Version]
- Bizzarri, M.; Dinicola, S.; Bevilacqua, A.; Cucina, A. Broad Spectrum Anticancer Activity of Myo-Inositol and Inositol Hexakisphosphate. Int. J. Endocrinol. 2016, 2016, 5616807. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; He, Y.; Wang, Y.-L.; Zhou, J.-G.; Chen, C. The efficacy and safety of inositol supplementation in preterm infants to prevent retinopathy of prematurity: A systematic review and meta-analysis. BMC Ophthalmol. 2019, 19, 135. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Study | Year | Evaluated INS | Study Characteristics | Main Results |
|---|---|---|---|---|
| Animal Studies—Deficiency | ||||
| Hayashi [36] | 1974 | MI | Rats fed with an MI-deficient diet or the same diet with the addition of 0.5% MI for up to five weeks. | Increased levels of liver TG, CE, and non-esterified fatty acids and concomitant increase in serum non-esterified fatty acids in the MI-deficient group. |
| Hayashi [37] | 1974 | MI | Rats fed with an MI-deficient diet or the same diet with the addition of 0.5% MI for one or two weeks. | Liver TG levels were increased in MI-deficient rats, especially in palmitic, palmitoleic, and oleic acids. |
| Thakur [38] | 2011 | PI | Zebrafish mutants incapable of PI synthesis. | Mutants exhibited hepatomegaly with microscopic NAFLD features with upregulated endoplasmic reticulum stress markers. |
| Animal Studies—Supplementation | ||||
| Katayama [39] | 1994 | MI | Rats fed with either corn starch or a high-sucrose diet, with or without MI, for 16–17 days. | Reduction in the increase of liver weight, total lipids, TG, and CE by MI in high-sucrose fed rats; reduction in serum TG increase in the same group. |
| Katayama [40] | 1997 | MI, sodium phytate | Rats fed with either corn starch or a high-sucrose diet, with or without the addition of MI or sodium phytate for 12–13 days. | MI and sodium phytate reduced liver enlargement and suppressed to normal levels liver TG and total lipids levels; reduced liver G6PD, ME, and FASN. |
| Geethan [41] | 2008 | Pinitol | Streptozotocin-induced diabetic rats treated with or without 100 mg/kg Pinitol for 30 days. | Pinitol reduced blood glucose and serum TG, free fatty acids, and CE; decreased TG and CE liver concentration; decreased the concentration of liver phospholipids and free fatty acids; increased HDL and reduced LDL. |
| Zhou [42] | 2008 | Pinitol | Rats fed with a high-fat diet for 8 weeks, with or without 0.1%, 1.0%, or 2.0% Pinitol, and induced hepatic injury by a single administration of GalN. | After GalN administration, Pinitol suppressed the increase in ALT and AST; attenuated liver CE increase; reduced TNFα levels; reduced lipid peroxidation; increased glutathione levels; increased liver catalase; Mn–SOD; GR activities. |
| Choi [43] | 2009 | Pinitol | Hamsters fed with a high-fat, high-cholesterol diet with or without 0.05% or 0.1% Pinitol for 10 weeks. | Pinitol reduced epididymal and perirenal white adipose tissue; reduced plasma total CE, non-HDL CE, glucose, and total-CE/HDL ratio; reduced liver TG and CE; lowered HMGR and ACAT activities; suppression of liver lipid accumulation and reduction in adipocyte size. |
| Sivakumar [44] | 2010 | Pinitol | Streptozotocin-induced diabetic rats treated with Pinitol, gliclazide, or neither for 30 days. | Both Pinitol and gliclazide reversed increase in blood glucose and glycosylated Hgb; reduced blood TNF-α, IL-6, and IL-1β; reduced liver peroxides and hydroperoxides; contrasted the diabetes-induced microscopic liver alterations normalizing the tissue architecture. |
| Shimada [45] | 2019 | MI | Rats fed with either a high-glucose or high-fructose diet, with or without MI 0.05% or 0.25% supplementation for 15 days. | MI dose-dependent reduction of liver TG content and expression levels of G6PD, ME1, FASN, ACCα, and S14 in fatty liver high-fructose induced rats; reduction in hepatic ChREBPβ expression; reduction in ChREBP binding to the ChoRE ChREBPβ and FASN genes. |
| Human Studies—Supplementation | ||||
| Lee [46] | 2019 | Pinitol | Double-blind RCT on 90 NAFLD patients taking Pinitol 600 mg, 1000 mg or PBO for 12 weeks | No significant between groups differences in liver fat content at 12 weeks; significant reduction in liver fat content in the 600 mg arm compared to its baseline. Pinitol significantly reduced AST levels at 12 weeks; reduced lipid peroxidation in terms of urinary MDA stability compared to PBO increased GPx. Pinitol reduced blood TG increase after postprandial high-fat formula compared to PBO. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pani, A.; Giossi, R.; Menichelli, D.; Fittipaldo, V.A.; Agnelli, F.; Inglese, E.; Romandini, A.; Roncato, R.; Pintaudi, B.; Del Sole, F.; et al. Inositol and Non-Alcoholic Fatty Liver Disease: A Systematic Review on Deficiencies and Supplementation. Nutrients 2020, 12, 3379. https://doi.org/10.3390/nu12113379
Pani A, Giossi R, Menichelli D, Fittipaldo VA, Agnelli F, Inglese E, Romandini A, Roncato R, Pintaudi B, Del Sole F, et al. Inositol and Non-Alcoholic Fatty Liver Disease: A Systematic Review on Deficiencies and Supplementation. Nutrients. 2020; 12(11):3379. https://doi.org/10.3390/nu12113379
Chicago/Turabian StylePani, Arianna, Riccardo Giossi, Danilo Menichelli, Veronica Andrea Fittipaldo, Francesca Agnelli, Elvira Inglese, Alessandra Romandini, Rossana Roncato, Basilio Pintaudi, Francesco Del Sole, and et al. 2020. "Inositol and Non-Alcoholic Fatty Liver Disease: A Systematic Review on Deficiencies and Supplementation" Nutrients 12, no. 11: 3379. https://doi.org/10.3390/nu12113379