
Abstract
Attention has often been considered to be a gateway to consciousness (Posner, Proceedings of the National Academy of Sciences of the United States of America, 91(16), 7398–7403, 1994). However, its relationship with conscious perception (CP) remains highly controversial. While theoretical models and experimental data support the role of attention in CP (Chica, Lasaponara, Lupiáñez, Doricchi, & Bartolomeo, NeuroImage, 51, 1205–1212, 2010; Dehaene, Changeux, Naccache, Sackur, & Sergent, Trends in Cognitive Sciences, 10, 204–211, 2006; Mack & Rock, Inattentional blindness, 1998), recent studies have claimed that at least some forms of attention—endogenous or top-down spatial attention—are neither sufficient nor necessary for CP (Koch & Tsuchiya, Trends in Cognitive Sciences, 11, 16–22, 2007). In the present experiments, we demonstrate the importance of exogenously triggered attention for the modulation of CP. Weak or null effects were instead observed when attention was triggered endogenously. Our data are discussed in the framework of recent neuropsychological models (Dehaene et al., Trends in Cognitive Sciences, 10, 204–211, 2006), postulating that activity within reverberating frontoparietal networks, as colocalized with spatial--orienting systems, is the brain correlate of consciously processed information.
Similar content being viewed by others


Introduction
Introspection suggests that when we attend to an object or part of a scene. we become conscious of it. However, the relation between spatial attention and conscious perception (CP) has proven intriguing and difficult to explore empirically. William James (1890) originally provided an influential definition of the interplay between attention and CP: “[Attention] is the taking possession of the mind, in clear and vivid form, of one out of several simultaneously possible objects or trains of thought.” This view led many to posit that spatial attention and CP are inextricably related (Bartolomeo, 2008; Chun & Marois, 2002; O'Regan & Noë, 2001; Posner, 1994). Some lines of evidence support this relationship. For example, in the inattentional blindness paradigm, salient changes in the features of visual stimuli are missed when unattended (Mack & Rock, 1998), even when the stimuli are presented at the fovea. Moreover, such a phenomenon is enhanced when the deployment of attention is challenged by increased levels of perceptual load (Lavie, 2006). Finally, strong evidence supporting the link between attention and CP comes from right-brain-damaged patients affected by spatial neglect, whose acquired inability in orienting attention toward the contralesional left space makes them unaware of stimuli presented within the neglected space (see Bartolomeo, 2007, for a recent review).
On the other hand, dissociations between some forms of attention and CP have also been recently reported (for reviews, see Koch & Tsuchiya, 2007; Lamme, 2003; see also Wyart & Tallon-Baudry, 2008). Crucially, all the previous studies in neurologically intact observers have investigated the relationship between endogenous forms of attention and CP. Altogether, the results from these studies maintain that under certain situations, endogenous attention is neither necessary nor sufficient for CP (Koch & Tsuchiya, 2007).Footnote 1
We have recently manipulated exogenous spatial attention and studied its effects on CP (Chica, Lasaponara, Lupiáñez, Doricchi, & Bartolomeo, 2010). Peripheral, spatially nonpredictive cues (which were well above the threshold of CP) preceded near-threshold targets. Our behavioral results revealed that exogenous attention increased the probability of conscious reports from the observers, who indicated having seen more targets at exogenously attended than at unattended locations. Event-related potentials demonstrated that an early-perceptual P100 component elicited by the cue correlated with the levels of CP of subsequently presented targets (Chica et al., 2010). This evidence strongly suggests that, unlike endogenous orienting, exogenous orienting might indeed be an important modulator of CP.
We hereby present a series of experiments, carried out in normal participants using near-threshold targets, in which our aim was to explore the role of endogenous versus exogenous attention in CP, as well as their potential interactions. We measured discrimination responses and CP on every trial and manipulated target contrast so that a proportion of the targets were not consciously reported. In five experiments, we manipulated how spatial attention was triggered, either endogenously (using central cues) or exogenously (using peripheral cues). We also studied the effects of the maintenance or removal of attention from exogenously attended locations (using peripheral predictive cues). In Experiments 1 and 2, attention was endogenously oriented using a central symbolic cue to a location in space before the target was presented (endogenously triggered and maintained attention). In Experiments 3 and 4, we used peripheral cues that were either nonpredictive (exogenously triggered attention and no endogenous maintenance or removal, as in Chica et al., 2010) or predictive of the appearance of the target at a given location. Cues could be predictive of either the same location of target appearance (i.e., predictive cues; exogenously triggered attention and endogenous maintenance) or the opposite location (i.e., counterpredictive cues; exogenously triggered attention and endogenous removal). Finally, in Experiment 5, we directly compared the effects of predictive central symbolic and peripheral cues on CP, because, in both cases, attention was maintained endogenously at the cued location, whereas the two conditions differed in how attention was triggered (endogenously or exogenously, respectively). In the last two experiments, we used signal detection theory (SDT) to explore the effects of spatial attention in perceptual sensitivity and decision biases.
Experiment 1
In this experiment, attention was triggered and maintained endogenously to a location where a near-threshold stimulus could appear. On the basis of previous findings (Kentridge, Nijboer, & Heywood, 2008; Koch & Tsuchiya, 2007; Wyart & Tallon-Baudry, 2008),Footnote 2 we expected that this form of attention would not increase the probability that the target would be consciously perceived.
Method
Participants
Twelve psychology students from the University of Granada took part in the experiment for course credit. Their ages ranged from 17 to 28 years (M = 21). All of the participants in this and the following experiments were naïve as to the purpose of the experiment and reported having normal or corrected-to-normal vision.
Apparatus and stimuli
E-Prime software was used to control the presentation of the stimuli, timing operations, and data collection (Schneider, Eschman, & Zuccolotto, 2002). The stimuli were presented on a gray background. Three black boxes (3.6° × 3.6°) were displayed, one at the center of the screen, and the other two centered 5.4° above and below it. The fixation point consisted of a black plus sign (3° × 3°) situated in the middle of the central box. The target consisted of a grating with a spatial frequency of 5 cycles per degree of visual angle and a diameter of 2.4°. The grating was tilted by 5° either to the left or to the right. Target contrast was manipulated before the experiment in order to adjust the percentage of consciously perceived targets between 60% and 75%. During practice trials, all the participants started with the high-contrast stimulus (Michelson contrast = 0.34), which was well above the threshold of CP. Every 16 trials, target contrast was automatically adjusted, so that when conscious detection levels proved higher than 75% correct, target contrast was decreased until an undetectable minimal contrast was reached (Michelson contrast = 0.009). Correspondingly, whenever performance was lower than 60% correct, target contrast was increased up to the maximum level (Michelson contrast = 0.34). There were 12 contrast levels between these maximum and minimum values, in which target contrast varied equidistantly. The experimental trials started when participants felt comfortable with the task and target contrast yielded ~75% seen targets. To ensure that factors such as practice or fatigue did not influence CP, the titration procedure operated every 16 trials during the whole experiment.
Procedure
Figure 1 displays the sequence and timing of a trial. The cues consisted of two letters: AR- from “arriba” (“up” in Spanish), AB- from “abajo” (“down”), and AA; the latter was a neutral cue indicating that the targets were to be presented with equal probability at the top and bottom locations. The up and down cues were predictive about the future location of the target on 75% of the target-present trials. Participants were informed about the predictive value of the cue, although they were not told about the exact number of trials on which the cue predicted the target location. They were encouraged to take this information into account in order to respond more accurately. The fixation point lasted for 515, 1,030, or 1,545 ms. Its duration was not fixed in order to avoid temporal preparation effects. Since central cues had to be processed and correctly interpreted in order to orient attention to its location, they were presented for 300 ms. There was a single interstimulus interval (ISI), which, on each trial, randomly took a value between 200 and 300 ms. This ensured that participants would have enough time to endogenously orient attention to the location indicated by the cue. The target was presented for 16 ms either at the upper or at the lower location, but never at the central location. Participants were asked to perform two responses consecutively, using target information. First, they were asked to discriminate the orientation of the grating (objective task) by pressing the “1” key on the numeric keypad of the keyboard if the lines were oriented to the left and the “2” key if they were oriented to the right. Participants responded with their right hand and were encouraged to respond on every trial as quickly and as accurately as possible. Even if they did not see the stimulus, they were told to try to guess their response. Second, they saw the question, “Did you see the stimulus?” (subjective task). This time, we encouraged participants to take their time to respond correctly and to report having seen the stimulus only when they were completely confident about having seen it. After every 16 trials, a message box appeared on the screen, indicating the percentage of correct rejections (trials on which the stimulus was not present and participants reported not having seen it). Participants were told to try to keep this percentage of correct rejections close to 100%. The experiment consisted of a total of 600 trials, 120 of which were target-absent trials. Valid trials were 3 times more likely than invalid and neutral trials (i.e., there were 3 valid trials for each invalid and each neutral trial).
Results
Participants were able to perform the task at a high level of accuracy concerning the presence or absence of the target in the subjective task. Their d’sFootnote 3 ranged from 1.14 to 2.93 (mean d’ = 2.25). χ² tests showed that the d’ value was different from zero for each participant (all ps < .001). The rate of false alarms was low (7%).
Reaction times on objective task
Reaction times (RTs) for correct responses shorter than 150 ms (5% of the trials) were considered anticipations and were eliminated from the RT analysis. We performed a repeated measures analysis of variance (ANOVA) on both correct and incorrect responses, with awareness (reported as seen vs. unseen targets) and validity (valid, invalid, and neutral trials) as factors. Only the main effect of awareness reached statistical significance, F(1, 11) = 15.41, MSE = 52,191, p < .01, indicating faster responses when participants reported having seen the targets. However, a putative effect of endogenous attention on the location indicated by the cue should be expected mostly when participants consciously perceived the target and correctly detected its orientation. We thus performed a separate one-way ANOVA for seen targets, which yielded no significant effect of validity on RTs, F < 1.
Mean accuracy on the objective task
Again, only the main effect of awareness proved significant, F(1, 11) = 26.12, MSE = 0.038, p < .05, because participants were more accurate when they reported having seen the targets. Performance was not different from 50% correct (i.e., chance level) when participants reported not having seen the target (M = .48), t = ˗1.30, p = .22, but it was above chance when participants reported having seen the target (M = .71), t = 4.32, p < .05. A separate ANOVA for seen targets indicated that participants were actually more accurate in discriminating targets preceded by valid and neutral cues than by invalidly cued targets, F(2, 22) = 4.39, MSE = 0.001, p < .05 (see Table 1).Footnote 4 This result indicates that endogenous cues were indeed effective in orienting attention toward the cued side.
Proportion of seen targets (subjective task)
Although endogenous orienting was effective in orienting participants’ attention to the location indicated by the cue, as it is demonstrated by the accuracy results for the objective task, it did not reliably modulate the proportion of consciously perceived targets at the valid, invalid, and neutral locations, F(2, 22) = 1.61, MSE = 0.001, p = .22 (see Table 1).
Discussion
The results of this experiment indicate that, unlike exogenous orienting (Chica et al., 2010), endogenously paying attention to the location indicated by a central symbolic cue does not significantly increase the proportion of consciously reported targets. This null result is in sharp contrast to the effect of endogenous attention on the objective task: Participants discriminated the orientation of the stimuli less accurately for invalid than for neutral and valid targets, which proves that they were actively using the cues to orient their attention endogenously, although CP was not enhanced.
The relatively inefficient influence of endogenous attention in CP is consistent with previous data obtained with a variety of different methods (see Kentridge et al., 2008, for a behavioral dissociation; see Wyart & Tallon-Baudry, 2008, for an electrophysiological data dissociation). However, the present findings seem, at first sight, inconsistent with a recently published article reporting that endogenously triggered attention increased contrast appearance in an objective task (Liu, Abrams, & Carrasco, et al. 2009). This discrepancy may result, among other causes, from differences in target saliency, which was set up well above the threshold for CP in Liu et al.’s study and at near-threshold levels for CP in the present study. Therefore, even though endogenous attention could enhance contrast perception for consciously perceived, suprathreshold targets (Liu et al., 2009), it may still produce a weaker or null modulation for near-threshold targets.
Experiment 2
To test the latter hypothesis, in this experiment, we used a paradigm similar to that applied in Experiment 1, with the difference that participants were presented this time with two blocks of trials. In the first block, all the stimuli were clearly presented above threshold. In the second block, the same near-threshold stimuli as those used in Experiment 1 were used. Furthermore, eye movements were controlled at all times in order to ensure that the results could not be accounted for by uncontrolled eye movements. We expected to replicate the finding that endogenous attention does not increase the percentage of consciously perceived targets in Block 2 (near-threshold targets), and eventually predicted, in agreement with prior reports (Liu et al., 2009), a modulation of the objective task in Block 1 (suprathreshold targets).
Method
Participants
A new sample of 14 naïve French participants took part in the experiment. Participants’ ages ranged from 19 to 33 years (M = 25). They were rewarded with 20€ for their participation.
Apparatus, stimuli, and procedure
Everything was the same as in Experiment 1, except that the cues were written in French: HA- from “haut” (“up” in French) and BA- from “bas” (“down”). No neutral cue was used in this experiment. The up and down cues were predictive of the future location of the target on 75% of the target-present trials. The experiment was composed of two blocks of trials. During the first block (300 trials), all the targets were high-contrast stimuli (the higher contrast used in Experiment 1; Michelson contrast = 0.34), well above the threshold for CP. During the second block of trials (300 trials), target contrast was automatically adjusted before the experiment and every 16 trials, as in the previous experiment. This manipulation ensured that about 25% of the targets were not consciously reported. The suprathreshold block was always performed first, to ensure that participants could capture the cue–target contingency and could orient their attention accordingly.
The stimuli were presented on an eyetracker screen (Tobii Technology AB, Danderyd, Sweden), 17 in. wide, 1,024 × 768 (16-ms refresh rate; temporal and spatial resolution of 50 Hz and 0.25°, respectively). The experiment consisted of a total of 600 trials. Each suprathreshold and near-threshold block was composed of 300 trials, 60 of which were no-target trials. Valid trials were 3 times more likely to occur than invalid trials.
Results
Participants were able to perform the task at a high level of accuracy concerning the presence or absence of the target in the subjective task. Their d’s ranged from 2.45 to 4.73 (mean d’ = 3.62) in the suprathreshold targets block and from 1.55 to 2.60 (mean d’ = 2.23) in the near-threshold targets block. χ² tests showed that the d’ value was statistically different from zero for each participant and each block of trials (all ps < .001). The rate of false alarms was 5.7% and 2.4% for the suprathreshold and near-threshold blocks of trials, respectively.Footnote 5
Two participants were eliminated from the analysis because they broke fixation on more than 30% of the trials in at least one of the blocks. For the remaining 12 participants, eye movements were very rare and occurred on only 3% of the trials in the suprathreshold block and 6% of the trials in the near-threshold block. All trials on which correct fixation was broken were eliminated from further analysis.
RTs on the objective task
RTs shorter than 150 ms (3.6% and 3.4% of correct trials for the suprathreshold and near-threshold blocks of trials, respectively) were eliminated from the RT analysis as anticipations. We performed a repeated measures ANOVA with block (suprathreshold and near-threshold targets) and validity (valid and invalid trials) for seen targets as factors. There was a main effect of block, because RTs were shorter for suprathreshold than for near-threshold targets, F(1, 11) = 6.08, MSE = 8,073, p = .031. The main effect of validity was also significant, F(1, 11) = 5.38, MSE = 1,010, p = .040, because responses were faster for valid trials than for invalid trials, consistent with attention being oriented toward the cued location. Importantly, this validity effect was similar for the suprathreshold and the near-threshold blocks (F < 1, for the interaction between validity and block; see Table 2).
Mean accuracy on the objective task
A similar analysis of the mean accuracy data showed that the only significant effect was the main effect of block, F(1, 11) = 36.34, MSE = 0.005, p < .001, with more accurate responses for suprathreshold than for near-threshold targets.
Proportion of seen targets (subjective task)
Only near-threshold targets were analyzed, because participants consciously perceived nearly all the suprathreshold targets. We performed a one-way ANOVA with the factor of validity. The results showed that endogenously attending to the centrally cued location did not increase the percentage of consciously perceived targets, F < 1.
Following Wilimzig, Tsuchiya, Fahle, Einhauser, and Koch (2008), and in order to directly compare the modulation observed in the objective and subjective tasks, we computed the mean Δz scores for the RT data recorded throughout the objective task (mean RTs for invalid minus valid trials) and the data for the subjective task (mean proportion of seen targets for valid minus invalid trials) in Experiments 1 and 2. Mean RT for each condition was divided by the standard deviation of the overall mean. Then each condition’s z-score was subtracted between conditions to compute Δz scores. Although a direct comparison of the scores on the objective and subjective tasks did not yield statistically significant differences (p = .72; mean Δz score for the objective task, 0.54; mean score for the subjective task, 0.37), t-tests revealed a significant effect of validity on the objective response (p = .04), but not on the subjective response (p = .38). This further indicates that endogenously triggered attention was relatively inefficient in increasing the probability of consciously reporting attended targets.
Discussion
The results from Experiment 2 confirmed that endogenous attention produced an effect in the objective task for suprathreshold targets, which is consistent with Liu et al.’s (2009) data. We also replicated Experiment 1’s main finding: When endogenous attention is endogenously triggered and maintained, participants are not more likely to consciously report the appearance of the target, in spite of attention speeding up responses or enhancing accuracy at the attended location.
On the basis of the spotlight metaphor of attention (Posner, Snyder, & Davidson, 1980), most researchers still conceive spatial attention as a single mechanism that can be oriented to locations in space either endogenously or exogenously. If these forms of attention depended on a single mechanism, the effects of endogenous and exogenous attention in the processing of stimuli should be qualitatively similar, differing only in quantitative aspects, such as the speed of the attentional movement or the duration of its effects (Müller & Rabbitt, 1989). However, recent research has clearly shown that endogenous and exogenous attention produce qualitatively different effects on the processing of information (Botta, Santangelo, Raffone, Lupiáñez, & Belardinelli, 2010; Funes, Lupiáñez, & Milliken, 2007; see Klein, 2004, for a review), can be deployed independently of each other (Berger, Henik, & Rafal, 2005; Chica, Lupiáñez, & Bartolomeo, 2006), and are implemented in partially distinct frontoparietal regions (Chica, Bartolomeo, & Valero-Cabré, 2011; Corbetta & Shulman, 2002). This accumulating evidence suggests that endogenous and exogenous attention consist of two separate attentional systems. We propose that exogenously triggered attention modulates conscious perception, whereas, as has been suggested in prior studies (Kentridge et al., 2008; Wyart & Tallon-Baudry, 2008) and in the present series of experiments, endogenously triggered attention does not seem to play an equivalent role.
However, the fact that endogenous attention was not proven to modulate CP in our experiments (while triggered using central symbolic cues) does not imply that it plays no role at all. As it was noted in the introduction, an important dissociation refers to how attention is triggered and how it is maintained. This distinction is crucial to understanding the effects of central and peripheral cuing. For example, it has been demonstrated that when attention is triggered and maintained using central cues, it does not induce the illusory line motion (Christie & Klein, 2005). However, maintaining or removing endogenous attention from a location where attention has been exogenously triggered (using a peripheral cue) does modulate the illusion, which is nevertheless generated only by exogenous peripheral cues (Chica, Charras, & Lupiáñez, 2008). Likewise, although Experiments 1 and 2 demonstrated that central cues do not modulate CP, we cannot rule out the potential for CP modulation if attention is initially triggered exogenously using a peripheral cue but is maintained endogenously through cue–target predictability.
Experiment 3
In order to test the latter hypothesis, in Experiments 3 and 4, we used peripheral cues that were either nonpredictive of the future location of the target (exogenously triggered attention, with no endogenous orienting component) or predictive of target appearance. Cues were predictive of targets appearing at the same location as the cue (exogenously triggered attention and endogenous maintenance) or at the opposite location (exogenously triggered attention and endogenous removal; see Chica et al., 2008, for a similar design). Experiment 4 was very similar to Experiment 3, but its design allowed us to measure perceptual sensitivity and response criterion for valid and invalid trials. Moreover, eye movements were monitored in order to avoid any possible explanations of our results in terms of overt, rather than covert, orienting.
Method
Participants
A new sample of 12 naïve participants from the University of La Sapienza, Rome, took part in the experiment. Participants’ ages ranged from 23 to 26 years (M = 25). They received course credits as a reward for their participation.
Apparatus, stimuli, and procedure
At variance with the previous experiments, which employed central symbolic cues, Experiment 3 used peripheral cues lasting for 50 ms. Cues consisted of a square surrounding either the upper or the lower placeholder marker. Two cue-to-target ISIs, varying randomly between 200 and 300 ms or between 900 and 1,000 ms, were used. No neutral cue was used in this experiment. After the objective grating tilt discrimination task, participants were required to report whether they had seen the target or not. At variance with the previous experiments, in which the same keys were always assigned to the “yes” and “no” responses, in Experiment 3, below the text string “Did you see the stimulus?” the words “yes” and “no” were presented either at the left or at the right of the fixation point. The location of the words varied on every trial, and participants were required to press the “1” button to choose the word presented on the left and the “2” button for the right-presented word. This was done to avoid response preparation before the subjective question was presented.
There were three blocks of trials. In the first block (nonpredictive cue), the peripheral cue was not informative about the future location of the target, and therefore, on 50% of the trials the cue was presented at the same location as the impending target (valid trial), and on the remaining 50% at the opposite location (invalid trial). In the other two blocks, the cue was informative about the future location of the target on 75% of the target-present trials. More specifically, in the predictive-cue block, the cue predicted that the target would appear at the same location (75% valid trials and 25% invalid trials), whereas in the counterpredictive-cue block, the cue predicted that the target would appear at the location opposite to the cue (25% valid trials and 75% invalid trials). The nonpredictive cue block was always presented first and consisted of 720 trials. The order of the predictive and counterpredictive cue blocks was counterbalanced between participants. Each block consisted of 1,200 trials. In each block, the target was absent on 240 trials.
Results
Participants reported the presence or absence of the target with good accuracy. Their d’s ranged from 1.32 to 3.09 (mean d’ = 2.40) in the nonpredictive block, from 1.05 to 2.79 (mean d’ = 2.02) in the predictive block, and from 1.53 to 3.39 (mean d’ = 2.25) in the counterpredictive block. χ² tests showed that the d’ value was different from zero for each participant and block of trials (all ps < .001). The rate of false alarms was low (6%, 9%, and 7% for nonpredictive, predictive, and counterpredictive cue blocks, respectively).
RTs on the objective task
RTs shorter than 150 ms were eliminated from the RT analysis as anticipations (1.06%, 1.66%, and 1.45% of correct trials for nonpredictive, predictive, and counterpredictive blocks, respectively). In this experiment and in Experiment 5, in which the stimulus onset asynchrony (SOA) was manipulated, there were more instances of excessively slow responses than in the previous experiments, because the time of target appearance was not constant. To reduce variance in the data, RTs longer than 1,500 ms were also eliminated as outliers (1.19%, 2.30%, and 1.60% of correct trials for nonpredictive, predictive, and counterpredictive blocks, respectively).
We performed a repeated measures ANOVA on seen targets, with the factors of block (nonpredictive, predictive, and counterpredictive), validity (valid and invalid), and SOA (short and long) (see Table 3). Validity interacted with SOA, F(1, 11) = 6.39, MSE = 1,575, p = .028, with a 27-ms facilitatory effect at the short SOA and a 6-ms trend toward inhibition (shorter RTs for invalid than for valid trials) at the longer SOA. Importantly, there was an interaction between block and validity, F(1, 11) = 8.06, MSE = 897, p = .002, demonstrating that participants were correctly orienting attention in relation to the informative value of the cue. As can be observed in Fig. 2, the expected results were observed. At the short SOA, a further ANOVA test revealed that the interaction between block and validity was significant, F(2, 22) = 7.62, MSE = 979, p = .003, indicating that there was a facilitatory effect for the nonpredictive and predictive cue blocks, which was not observed for the counterpredictive block. At the long SOA, however, the interaction between validity and block was marginally significant, F(2, 22) = 2.82, MSE = 410, p = .082, with a tendency for faster responses for invalid than for valid trials in the nonpredictive and the counterpredictive cue blocks, but not in the predictive block.

Sequence of events on a given trial. An example of the cues and targets used in Experiments 1 and 2 is shown

The left panel shows the mean reaction times (RTs) for the objective task, and the right panel shows the proportions of seen targets in the subjective task, for each validity condition as a function of block and stimulus onset asynchrony (SOA) in Experiment 3
Mean accuracy on the objective task
We performed a repeated measures ANOVA on the mean accuracy rates, with block (nonpredictive, predictive, and counterpredictive), validity (valid and invalid), and SOA (short and long) for seen targets as factors. It revealed that only the main effects of validity and SOA were significant, F(1, 11) = 6.14, MSE = 0.003, p = .031, and F(1, 11) = 5.57, MSE = 0.003, p = .038, respectively, with more accurate responses for valid than for invalid trials and for short than for long SOAs.
Proportion of seen targets (subjective task)
We performed a repeated measures ANOVA on the mean proportion of consciously reported targets, with the factors of block (nonpredictive, predictive, and counterpredictive), validity (valid and invalid), and SOA (short and long). This analysis revealed a main effect of SOA, F(1, 11) = 7.83, MSE = 0.014, p = .017, indicating that participants reported having seen more targets at the shorter SOA than at the longer one. The interaction between block, validity, and SOA was significant, F(1, 11) = 3.61, MSE = 0.001, p = .044 (see Table 3). In order to better understand this interaction, we performed two further ANOVAs comparing the nonpredictive and predictive cue blocks and the nonpredictive and counterpredictive cue blocks, respectively. When the nonpredictive and predictive cue blocks were compared, there was only a marginally significant main effect of validity, F(1, 11) = 4.08, MSE = 0.029, p = .068, with more consciously reported targets for valid than for invalid trials in both blocks. However, when the nonpredictive and the counterpredictive cue blocks were compared, block interacted with validity, F(1, 11) = 5.35, MSE = 0.003, p = .041, because more targets were reported for valid than for invalid trials in the nonpredictive cue block, but not in the counterpredictive cue block (see Fig. 2). This result indicates that endogenously removing attention from the location of the cue eliminated the benefit observed for valid over invalid trials when endogenous attention remained at that cued location.
Discussion
The results of Experiment 3 are especially relevant since they show that when attention is exogenously triggered using peripheral cues (either predictive or nonpredictive), a modulation of CP may follow. However, when attention is endogenously removed from peripheral cues, there are no effects on CP, an observation that is consistent with the results observed with central endogenous cues in Experiments 1 and 2. It could have been interesting to measure d’ and beta to determine whether the attentional modulations of CP were due to changes in perceptual sensitivity or response biases. This was not possible with the present design, since when no target was presented, false alarms could not be classified as being valid or invalid.
Experiment 4
To address this issue, the present experiment was designed, which allowed us to assign false alarms to valid or invalid trials and, thus, compute SDT analyses. Moreover, eye movements were also monitored to avoid any potential bias caused by overt, rather than covert, attentional orienting.
Method
Participants
A new sample of 16 naïve participants from Paris, France, took part in the experiment. Participants’ ages ranged from 18 to 29 years (M = 24). They were rewarded with 20€ for their participation.
Apparatus, stimuli, and procedure
Everything was performed as in Experiment 3, except that eye movements were monitored using the same device as that in Experiment 2 and except for other changes stated below. The markers (6° width × 5.5° height) were horizontally aligned. The fixation point (0.5 × 0.5°) was presented for a time ranging from 500 to 1,000 ms. The peripheral cue consisted of a black dot (0.5° diameter) presented in the upper external part of one of the boxes. In contrast to Experiment 3, there was a single cue–target ISI of 200 ms. The target was a Gabor stimulus (4 cycles/deg spatial frequency, 3° in diameter, SD of 0.3°), with a maximum and minimum Michelson contrast of 0.56 and 0.02, respectively. There were 40 stimuli between these maximal and minimum values. In order to be able to classify false alarms as valid (same location as the previous cue) or invalid (opposite location than the previous cue), during the subjective task, we presented participants with two arrow-like stimuli, one below and the other one above the fixation point (>>> or <<<). We provided participants with three keys (which they should press with their left hand): an upper key (“d”), a lower key (“c”), and the space bar. The upper key always corresponded to the arrow presented in the upper part of the fixation point, while the lower key was associated with the arrow presented in the lower part of the fixation point. Participants were asked to report, as accurately as possible, whether they had seen the target or not. If they had not, they were required to press the space bar. If they had seen the target, they were asked to indicate its location on the screen, left or right. This procedure allowed us to know whether a false alarm corresponded to the same location as the previous cue (valid) or to the opposite location (invalid).
The titration procedure was similar to that in the previous experiments (i.e., it was done before the experimental trials and was adjusted every 20 trials during the experiment) and ensured that participants reported having seen the targets on at least 67% of the trials. In this experiment, accuracy of the objective response was also titrated so that the correct discrimination performance was between 65% and 85% (Gabor grating tilt orientation ranging from 1° to 10°). In this and the following experiment participants were not instructed to keep the percentage of false alarms close to 0%, but they were informed about the percentage of false alarms committed if it was larger than 55% on the last 20 trials. As in Experiment 3, there were three blocks of trials: nonpredictive, predictive, and counterpredictive. Each block consisted of a total of 400 trials; in each block, the target was absent on 80 trials. In the predictive block, valid trials were 3 times more likely than invalid trials, while in the counterpredictive cue block, invalid trials were 3 times more likely than valid trials.
Results
Three participants were eliminated from the analyses because they broke fixation on more than 50% of the trials. Participants were able to perform the subjective task at a high level of accuracy regarding the presence or absence of the target. Their d’s ranged from 1.89 to 2.71 (mean d’ = 2.16) in the nonpredictive block, from 1.34 to 2.64 (mean d’ = 2.02) in the predictive cue block, and from 1.82 to 2.88 (mean d’ = 2.88) in the counterpredictive cue block. χ² tests showed that the d’ value was different from zero for each participant and block of trials (all ps < .001). The rate of false alarms for nonpredictive, predictive, and counterpredictive cue blocks was 4.77%, 4.53%, and 2.79%, respectively. Participants broke fixation on 8%, 13%, and 7% of the trials in the nonpredictive, predictive, and counterpredictive cue blocks, respectively. These trials were eliminated from further analyses.
RTs on the objective task
RTs shorter than 150 ms (6.8%, 2.0%, and 2.3% of correct trials, for nonpredictive, predictive, and counterpredictive blocks, respectively) were considered anticipations and were eliminated from the RT analysis. We performed a repeated measures ANOVA with the factors of block (nonpredictive, predictive, and counterpredictive) and validity (valid and invalid) for seen targets (see Table 4). As in Experiment 3, there was an interaction between block and validity, F(2, 24) = 11.61, MSE = 7,739, p < .001, demonstrating that participants were correctly orienting attention according to the informative value of the cue (see Fig. 3 and Table 4). RTs were shorter for valid than for invalid trials in the nonpredictive, F(1, 12) = 10.83, MSE = 1,580, p = .006, and predictive, F(1, 12) = 16.66, MSE = 10,723, p = .001, cue blocks, while the effect was reversed in the counterpredictive cue block, F(1, 12) = 7.21, MSE = 4,330, p = .020.

The left panel shows the mean reaction times (RTs) for the objective task, and the right panel shows the proportions of seen targets in the subjective task, for each validity condition as a function of block in Experiment 4
Mean accuracy on the objective task
We performed a repeated measures ANOVA on the mean accuracy rates, with block (nonpredictive, predictive, and counterpredictive), and validity (valid and invalid) for seen targets as factors, but none of the main effects or interactions reached statistical significance.
Proportion of seen targets (subjective task) and SDT analyses
We performed a repeated measures ANOVA on the mean proportion of consciously reported targets, with the factors of block (nonpredictive, predictive, and counterpredictive) and validity (valid and invalid). Replicating Experiment 3, we found a significant interaction between block and validity, F(2, 24) = 13.55, MSE = 0.015, p < .001, indicating that participants reported seeing more targets at valid than at invalid locations in the nonpredictive, F(1, 12) = 35.43, MSE = 0.003, p < .001, and predictive, F(1, 12) = 39.15, MSE = 0.017, p < .001 blocks, but not in the counterpredictive cue block, F < 1.
The analysis of d’ showed an interaction between block and validity, F(2, 24) = 5.36, MSE = 0.327, p = .012, demonstrating that making the cue spatially predictive produced an increase in d’ for valid, as compared with invalid, trials, F(1, 12) = 8.87, MSE = 0.44, p = .011, whereas it did not reach significance when the cue was spatially nonpredictive, F(1, 12) = 2.06, MSE = 0.18, p = .176, or counterpredictive (F < 1; see Table 4). There was also an interaction between block and validity in beta values, F(2, 24) = 9.89, MSE = 25.90, p < .001, indicating that response criterion was more relaxed for valid than for invalid trials in the nonpredictive, F(1, 12) = 16.88, MSE = 42.29, p = .001, and predictive, F(1, 12) = 10.64, MSE = 44.03, p = .007, blocks, while there were no significant differences for counterpredictive cues, F < 1.
Discussion
The results of Experiments 3 and 4 are important because they show that when attention is triggered exogenously, using peripheral cues (either predictive or nonpredictive), a modulation of CP may occur. However, endogenous attentional orienting to a noncued location (in the case of counterpredictive cues) yielded no effects on CP, consistent with the results observed with central cues in Experiments 1 and 2.
Changes in perceptual sensitivity are additionally observed when attention is both triggered exogenously and maintained endogenously (i.e., predictive peripheral cues). Importantly, endogenously attending to an uncued location (in the case of counterpredictive cues) did not modulate CP, which suggests that the sequence “exogenous attentional capture and endogenous maintenance of attention at the same location” sets up the best conditions for modulating CP. These results are overall consistent with the findings reported by Prinzmetal, McCool, and Park (2005), who proposed that while endogenous attention enhances perceptual representations, exogenous attention mostly affects decision stages. Our results show that although exogenously triggered attention produces changes in response criterion, additional changes in perceptual sensitivity are observed when endogenous attention is maintained at the cued location. What is important to note is that in order to modulate CP, spatial attention needs to be triggered exogenously, producing its maximal effects when attention is endogenously maintained at the cued location. Endogenously triggered attention (either elicited using central symbolic cues, as in Experiments 1 and 2, or oriented to a noncued location, as in Experiments 3 and 4) was much less effective in modulating CP.
In a last attempt to directly compare the effects of endogenously versus exogenously triggered attention in conscious perception, we ran Experiment 5, in which attention was endogenously maintained at a peripheral location but was triggered either endogenously (using a central symbolic cue) or exogenously (using a peripheral cue). On the basis of our previous data, we predicted that, despite similar endogenous maintenance of attention, exogenously triggered attention should produce larger effects on CP than endogenously triggered attention.
Experiment 5
In this experiment, the effects of central and peripheral predictive cues were compared while using the method of Experiment 4 to measure d’ and beta. Two SOAs were used in order to maximize the effects of both types of cues: a short SOA, at which the peripheral cue should produce its larger effects, and a long SOA, at which the central cue should produce its maximal effects.
Participants
A new sample of 8 naïve participants from Paris, France, took part in the experiment. Participants’ ages ranged from 22 to 36 years (M = 27). They were rewarded with 40€ for their participation in the two sessions of the experiment.
Apparatus, stimuli, and procedure
Everything was the same as in Experiment 4, except for the following. Two cue types (central and peripheral) were tested in separate sessions, with their order of presentation counterbalanced across sessions. For both cue types, the same circle was used as a cue. The central cue consisted of a circle that was either red or green in color (0.5° diameter), overlapping the fixation point, and presented during 50 ms. As in Funes et al. (2007), for half of the participants the red color meant target appearance more likely on the right, and green signaled target appearance more likely on the left. For the other half of the participants, this mapping was reversed. The use of colors to code space ensured that attention was symbolically driven. The peripheral cue was exactly the same as in Experiment 4. There were two ISIs between cue and target appearance: 200 and 1,500 ms. Each session consisted of a total of 800 trials, 160 of which were catch trials. Valid trials were 3 times more likely than invalid trials.
Results
Participants were able to perform the subjective task at a high level of accuracy regarding the presence or absence of the target. Their d’ ranged from 2.34 to 3.18 (mean d’ = 2.83) in the central cue condition and from 2.43 to 2.98 (mean d’ = 2.74) in the peripheral cue condition. χ² tests showed that the d’ value was different from zero for each participant and block of trials (all ps < .001). The rate of false alarms was 0.32% and 0.26% for the central and peripheral cue, respectively. Participants broke fixation on 11.48% of the trials in the central cue condition and on 9.58% of the trials in the peripheral cue condition. These trials were eliminated from further analyses.
RTs on the objective task
RTs shorter than 150 ms or longer than 1,500 ms (3.90% and 3.00% of correct trials for the central and peripheral cues, respectively) were considered anticipations or excessively slow responses and were thus eliminated from the RT analysis. We performed a repeated measures ANOVA with the factors of cue type (central and peripheral), validity (valid and invalid), and SOA (250 and 1,550 ms). The interaction between validity and SOA was significant, F(1, 7) = 10.53, MSE = 1,092, p = .014, the validity effect being significant only at the short SOA, F(1, 7) = 14.84, MSE = 1,328, p = .006. Importantly, the validity effect at this SOA was identical for central cues (44 ms) and for peripheral cues (54 ms; interaction between cue type, validity, and SOA, F < 1; see Fig. 4 and Table 5). This result confirms that participants were orienting attention effectively and in a similar way for either type of cue.

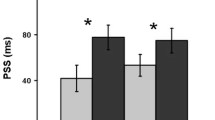
The left panel shows the mean reaction times (RTs) for the objective task, and the right panel shows the proportions of seen targets in the subjective task, for each validity condition as a function of cue type and stimulus onset asynchrony (SOA) in Experiment 5
Mean accuracy objective task
The ANOVA on the mean accuracy rates demonstrated a significant main effect of validity, F(1, 7) = 5.66, MSE = 0.004, p = .049, that did not depend on SOA, F(1, 7) = 1.99, MSE = 0.004, p = .20, or on cue type, F < 1.
Proportion of seen targets (subjective task) and SDT analyses
The ANOVA on the mean proportion of consciously reported targets revealed a main effect of SOA, F(1, 7) = 11.78, MSE = 0.069, p = .011, and an interaction between cue type, validity, and SOA, F(1, 7) = 14.36, MSE = 0.001, p = .007. Planned comparisons revealed that at the short SOA, validity effects were larger for peripheral cues than for central cues, F(1, 7) = 10.18, MSE = 0.001, p = .015. Participants perceived 13% more targets at the exogenously cued versus uncued location, but only 4% more targets at the endogenously attended versus nonattended location. At the long SOA, validity effects were slightly larger for the central than for the peripheral cue, F(1, 7) = 6.79, MSE = 0.001, p = .035, although this time the effect was smaller in size; participants perceived 4% more targets at the endogenously attended than at the nonattended location and only 1% more targets at the exogenously cued than at the uncued location.
We directly compared the validity effects of either cue type at the SOA at which its effects were maximal (short SOA for peripheral cues and long SOA for central cues). On the basis of the results of the previous experiments, we hypothesized that validity effects on CP would be larger for peripheral than for central cues. A one tailed t-test confirmed this hypothesis, t(1, 7) = 2.41, p = .047. It is important to note that in the central cue condition, although validity effects for the objective task decreased at the longer SOA, in the subjective task participants significantly reported seeing more targets for valid than for invalid conditions, the effect being identical at the long and short SOAs, t(1, 7) = 0.72, p = .49. Therefore, the effect of central cues on CP, although small, was independent of the SOA. This renders unlikely the possibility that using a different SOA would have increased the effects of endogenous attention on CP, since 1,550 ms seems a long enough SOA to observe endogenous effects (Funes et al., 2007; Müller & Rabbitt, 1989).
The d’ analysis revealed main effects of validity, F(1, 7) = 8.68, MSE = 0.12, p = .021, and SOA, F(1, 7) = 11.30, MSE = 0.58, p = .012, with better perceptual sensitivity for valid than for invalid trials and for short than for long SOAs. The interaction between cue type and validity was marginally significant, F(1, 7) = 4.47, MSE = 0.032, p = .072, suggesting a tendency for larger validity effects on d’ for peripheral than for central cues. Although the interaction between cue type, validity, and SOA did not reach significance, F(1, 7) = 2.18, MSE = 0.046, p = .183, the pattern of results was very similar to that obtained for the subjective reports (see Table 5). At the short SOA, validity effects tended to be larger for peripheral than for central cues, F(1, 7) = 4.60, MSE = 0.052, p = .069; at the long SOA, validity effects were similar for both types of cues, F < 1. In the beta analysis, none of the main effects or interactions reached significance.
Discussion
The results of Experiment 5 further demonstrate that endogenously triggered and maintained attention produces a weak influence on CP, as compared with exogenously triggered and endogenously maintained attention, which proved stronger and more replicable. This result was found for central versus peripheral cue conditions that were equated on cue predictability and demonstrated statistically similar attentional-orienting effects on the objective task. Moreover, the results of Experiment 5 confirm with a longer SOA those observed in Experiments 1 and 2. This result is reassuring because it goes against the possible concern that the lack of modulation of endogenously triggered attention on CP depended on the use of SOAs too short to endogenously orient attention to the indicated location.
In Experiments 3 and 5, in which the SOA was manipulated, participants reported consciously perceiving more targets at the short than at the long SOA, indicating that this variable had the ability to modulate subjective responses and perceptual sensitivity. This result is consistent with recent data from our group demonstrating that phasic alerting improves conscious perception and perceptual sensitivity (Kusnir, Chica, Mitsumasu, & Bartolomeo, 2011), and with results from the temporal-orienting literature showing that perceptual sensitivity is increased when targets appear at the expected moment in time (Correa, Lupiáñez, & Tudela, 2005; Rolke & Hofmann, 2007).
General discussion
Taken together, the five experiments of this study suggest that endogenously triggered attention is relatively inefficient at modulating CP for near-threshold stimuli. This result is consistent with those in previous literature claiming that some forms of attention are neither necessary nor sufficient for CP (Kentridge, Heywood, & Weiskrantz, 1999, 2004; Kentridge et al., 2008; Koch & Tsuchiya, 2007; Wyart & Tallon-Baudry, 2008). However, recent data from our group strongly indicate that, contrary to endogenous attention, exogenous orienting does seem to play an important role in CP (Chica et al., 2010). Indeed, Experiments 3, 4, and 5 show that CP is increased at exogenously triggered locations, while maintaining or removing endogenous attention from the exogenously attended location can also influence CP (Experiments 3 and 4). Therefore, it can be concluded that although endogenously triggered attention does not produce strong modulations of CP (at least within the experimental paradigms used in this study), it has the potential to interact with the effects of exogenous attentional orienting.
The present design employed stimuli that were not consciously perceived all the time. Following Dehaene, Changeux, Naccache, Sackur, and Sergent’s (2006) taxonomy, these stimuli could be considered as preconscious, and their access to consciousness may depend on the functional state of frontoparietal networks before or during stimulus presentation. Preparatory activity in frontoparietal cortices (Hopfinger, Buonocore, & Mangun, 2000) may thus influence CP for a given stimulus. This hypothesis is consistent with recently published evidence (Liu et al., 2009) demonstrating that endogenous attention can increase contrast perception when targets are presented well above the threshold for CP. However, our data indicate that endogenous spatial attention might not be the key factor for activating the frontoparietal networks claimed to be necessary for CP. This does not mean that such process is not required at all, but that it is at least insufficient to produce the top-down amplification that might precede consciousness. Exogenous attention, on the other hand, has been proven to be a powerful modulator of CP. This system might influence the activity of frontoparietal networks in such a way as to boost the feed-forward information necessary for the CP of ensuing targets (Fahrenfort, Scholte, & Lamme, 2008). The spatial coincidence between valid peripheral cues and correctly reported targets suggests that cues may act as “spatial anchors,” or “object files,” for attention to amplify objects subsequently appearing at that spatial location and prioritize them for access to CP.
Our subjective measure of CP depended on verbal reports. Drawing on the time-honored distinction between direct and reflective forms of consciousness (see, e.g., Merleau-Ponty, 1942), one could maintain that, although an appropriate verbalization can be considered to be a reliable indicator of conscious processing (Merikle, Smilek, & Eastwood, 2001), the converse is not necessarily true; lack of verbalization might simply indicate lack of reflective consciousness with normal direct consciousness (Bartolomeo & Dalla Barba, 2002). M. Snodgrass, Kalaida, and Winer (2009) interpreted this distinction in terms of an internal signal detection process. Reflective consciousness can access the contents of direct consciousness if they are both sufficiently clear to produce reasonable confidence in the perception and deemed task relevant. The present evidence (Experiment 4) in favor of a criterion shift with validly cued targets by peripheral cues seems consistent with this position, in the sense that valid cues might have increased participants’ confidence on their perception and, thus, shifted their criterion to more liberal levels.
Nonetheless, a general concern with the use of near-threshold stimuli, which makes the interpretation of those paradigms complex, is the fate of unseen stimuli.Footnote 6 If directing attention away from a stimulus causes failures in eliciting CP and influences visual behavior, one could wonder whether the unattended stimulus is having any effect on the attentional brain systems. In the present study, we cannot provide direct evidence on whether or not the stimulus appearing at a location remote from the direction of exogenous attention is or is not being processed at all. Nonetheless, we can draw supporting data from our own ERP recordings (Chica et al., 2010), whereby, replicating previous evidence (Sergent, Baillet, & Dehaene, 2005), we demonstrated that the evoked signals elicited by seen and unseen targets were very similar until around 300-ms post-stimulus-onset. Therefore, visual processing of seen and unseen stimuli would be similar up to the extrastriate visual areas (as indicated by the P100 component), while only seen targets elicit a P300 component, distributed throughout frontoparietal electrodes. These results strongly suggest that in our paradigm, unseen targets are visually computed at early stages of processing but might fail to reach CP.
How can the differences between exogenous and endogenous orienting in CP be accounted for? One possibility is that peripheral cues might initiate some activity at the corresponding spatial location (they might, for example, open an "object file"; Lupiáñez, 2010; Milliken, Tipper, Houghton, & Lupiáñez, 2000), such that targets subsequently presented at that location have a priority access to CP. Nevertheless, the extent to which this “object file” representation is used for further performance might depend on its ability to predict target appearance at a similar location; such piece of information might be integrated within the same representation. A complementary explanation might be related to the role of phasic alerting and its interactions with spatial orienting. We have demonstrated that phasic auditory alerting improves conscious perception and perceptual sensitivity (Kusnir et al., 2011). In Experiments 3 and 5 of the present study, we also found reliable effects of SOA, because more targets were consciously perceived at the short than at the long SOA. Thus, enhanced perceptual sensitivity at short SOAs might result from phasic alerting provided by the visual cue. Given that peripheral cues produce their larger effects at short SOAs, phasic alerting and spatial orienting could have additive effects. Central symbolic cues also produce phasic alerting, but their interpretation takes a longer time. Therefore, once attention is oriented to the cued location, alerting is not maximal, and only the isolated effects of spatial orienting, instead of their summation, are observed.
Our results might instead seem inconsistent with the conclusions of Woodman and Luck (2003). These authors used an “object substitution masking” paradigm to explore the role of attention in CP. In this paradigm, an object presented in a crowded environment is masked by the presentation of small objects surrounding it; when the mask offsets some time after the display onset, the masked object is not consciously perceived. In Woodman and Luck’s study, the N2pc ERP componentFootnote 7 was used to index the orienting of attention to the target. Their results showed that the N2pc was elicited both when the target was consciously perceived and when it was not, leading the authors to conclude that attention and CP are two independent processes and that the orienting of attention did not intrinsically produce conscious reports. However, as was noted by the authors, the N2pc ended earlier when the targets were not consciously perceived. If N2pc is a correlate of exogenous orienting of attention, these results can be taken as evidence in favor of the idea that exogenous attention is an important modulator of CP. The fact the N2pc ended earlier when the stimulus was not consciously reported might indicate that even if the target produced an exogenous attentional capture, the corresponding frontoparietal activation was unable to maintain the exogenous capture of attention long enough to trigger the necessary reverberation of information required for CP.Footnote 8 This is consistent with our proposal that exogenous attention is an important modulator of CP, although it may not be sufficient. Exogenously attended information is not always consciously perceived (Kentridge et al., 1999; McCormick, 1997; Mele, Savazzi, Marzi, & Berlucchi, 2008), perhaps because it does not always lead to the sustained frontoparietal activity that might be necessary for CP (Dehaene et al., 2006; Dehaene & Naccache, 2001). Neural information needs to reverberate in these networks (Fahrenfort, Scholte, & Lamme, 2007), and CP might be impaired if the system is not correctly prepared at the time information is being presented, or when brain damage interrupts the proper communication within and between these networks (Bartolomeo, Thiebaut de Schotten, & Doricchi, 2007). On the basis of the present study, we hypothesize that making the peripheral cues predictive might allow sustained reverberation on the attentional networks and thus increase the probability of CP.
In brain-damaged patients, CP might be impaired as a consequence of direct damage to areas or disconnection of sites within the brain networks including sensory primary areas and frontoparietal regions. In the blindsight patient G.Y., afflicted by a selective destruction of left primary visual cortex, effects of exogenous attention have been observed in the absence of CP (Kentridge et al., 1999). We propose that when the visual areas are disconnected from frontoparietal networks, CP will not emerge, because the visual input will be unable to undergo further processing. Even if an exogenous attentional capture occurs, information will not be able to reverberate within the frontoparietal network, thereby failing to produce CP. It is also tempting to relate the profound unawareness of contralesional stimuli demonstrated by patients to visual neglect with their prominent impairment in exogenous orienting. Importantly, neglect patients’ lesions typically involve the right parietal lobe (Mort et al., 2003) and its connections with the prefrontal cortex (Bartolomeo et al., 2007). Activity of such frontoparietal networks has been implicated both in orienting of attention (Buschman & Miller, 2007; Corbetta & Shulman, 2002; Nobre, 2001) and in consciousness (Dehaene et al., 2006). Neglect patients present profound deficits in exogenous, rather than endogenous, orienting, an observation that is consistent with the findings reported in this article about the special role of exogenous, rather than endogenous, orienting in CP.
Thus far, we have thoroughly empirically explored the type of attentional orienting necessary to influence CP by means of behavioral tasks, and we have related it to specific neuroanatomical hypotheses. We have developed and optimized a series of behavioral paradigms sensitive to the effects of cuing and their influence on conscious visual processes. Future research will be directed at further exploring the neural basis and temporal correlates of such processes by means of neuroimaging techniques such as ERPs, fMRI, and TMS. Such information might prove crucial to exploring ways to more efficiently “ignite” the workspace delimited between networks involved in sensory consciousness (Dehaene et al., 2006) and manipulate it at will for investigational or clinical purposes.
Notes
As was noted by Chica et al. (2010), Wyart and Tallon-Baudry (2008) used a central cue that indicated with a high probability where a near-threshold target would be presented. Their participants reported seeing more targets at valid than at invalid locations. However, the cue used by Wyart and Tallon-Baudry was not symbolic. It consisted of an arrow pointing to one of the possible locations where the target would be presented. These arrow cues have been demonstrated to produce an effective orienting of attention, even when they are not predictive about the future location of the target (Doricchi, Macci, Silvetti, & Macaluso, 2010; Tipples, 2002), suggesting that they trigger attention exogenously to their location. Thus, it is uncertain which forms of attentional orienting were modulated in Wyart and Tallon-Baudry’s study.
Zero false alarm rates were corrected in this and the following experiments using the following equation proposed by Snodgrass and Corwin (1988): FA = (FA + 0,5) / (FA + CR + 1,0).
Given that validity effects are expected mostly for seen targets, in order to simplify the analyses in the next experiments, only seen targets performance will be analyzed for the objective task.
As was expected given our manipulation, participants reported seeing more targets in the suprathreshold (97%) than in the near-threshold block (58%), p < .001. d′ was larger for suprathreshold than for near-threshold stimuli, p < .001, although more false alarms were also committed for suprathreshold than for near-threshold stimuli, p = .015. In the present study, near-threshold and suprathreshold targets refer to the likelihood that targets will be consciously acknowledged. Thus, the critical difference between the near-threshold and suprathreshold conditions lies in participants’ adjustment of their criteria for making positive reports.
We would like to thank Robert Kentridge for introducing this issue during the review process.
N2pc corresponds to N200 observed at parietal sites; it is uncertain whether it reflects exogenous attention, endogenous attention, or a combination of both.
In our experiments, participants were not unconscious of all the targets presented at the invalid (unattended) location. A strong version of our proposal would predict that because attention is oriented to the location opposite to the target on these trials, participants should always fail to report invalidly cued targets. However, even though attention is oriented to the location of the cue, the probability that an invalidly cued target attracts attention is low but not null. Thus, CP might have occurred for those invalidly cued targets that captured participants’ attention.
References
Bartolomeo, P. (2007). Visual neglect. Current Opinion in Neurology, 20, 381–386.
Bartolomeo, P. (2008). Varieties of attention and of consciousness: Evidence from neuropsychology. Psyche, 14(1)
Bartolomeo, P., & Dalla Barba, G. (2002). Varieties of consciousness (commentary on Perruchet and Vinter: The self-organizing consciousness). The Behavioral and Brain Sciences, 25, 331–332.
Bartolomeo, P., Thiebaut de Schotten, M., & Doricchi, F. (2007). Left unilateral neglect as a disconnection syndrome. Cerebral Cortex, 17, 2479–2490.
Berger, A., Henik, A., & Rafal, R. (2005). Competition between endogenous and exogenous orienting of visual attention. Journal of Experimental Psychology: General, 134, 207–221.
Botta, F., Santangelo, V., Raffone, A., Lupiáñez, J., & Belardinelli, M. O. (2010). Exogenous and endogenous spatial attention effects on visuospatial working memory. The Quarterly Journal of Experimental Psychology, 63, 1–13.
Buschman, T. J., & Miller, E. K. (2007). Top-down versus bottom-up control of attention in the prefrontal and posterior parietal cortices. Science, 315, 1860–1862.
Chica, A. B., Bartolomeo, P., & Valero-Cabré, A. (2011). Dorsal and ventral parietal contributions to spatial orienting in the human brain. Journal of Neuroscience.
Chica, A. B., Charras, P., & Lupiáñez, J. (2008). Endogenous attention and illusory line motion depend on task set. Vision Research, 48, 2251–2259.
Chica, A. B., Lasaponara, S., Lupiáñez, J., Doricchi, F., & Bartolomeo, P. (2010). Exogenous attention can capture perceptual consciousness: ERP and behavioural evidence. Neuroimage, 51, 1205–1212.
Chica, A. B., Lupiáñez, J., & Bartolomeo, P. (2006). Dissociating inhibition of return from the endogenous orienting of spatial attention: Evidence from detection and discrimination tasks. Cognitive Neuropsychology, 23, 1015–1034.
Christie, J., & Klein, R. M. (2005). Does attention cause illusory line motion? Perception & Psychophysics, 67, 1032–1043.
Chun, M. M., & Marois, R. (2002). The dark side of visual attention. Current Opinion in Neurobiology, 12, 184–189.
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed and stimulus-driven attention in the brain. Nature Reviews. Neuroscience, 3, 201–215.
Correa, A., Lupiáñez, J., & Tudela, P. (2005). Attentional preparation based on temporal expectancy modulates processing at the perceptual level. Psychonomic Bulletin & Review, 12, 328–334.
Dehaene, S., Changeux, J. P., Naccache, L., Sackur, J., & Sergent, C. (2006). Conscious, preconscious, and subliminal processing: A testable taxonomy. Trends in Cognitive Sciences, 10, 204–211.
Dehaene, S., & Naccache, L. (2001). Towards a cognitive neuroscience of consciousness: Basic evidence and a workspace framework. Cognition, 79, 1–37.
Doricchi, F., Macci, E., Silvetti, M., & Macaluso, E. (2010). Neural correlates of the spatial and expectancy components of endogenous and stimulus-driven orienting of attention in the posner task. Cerebral Cortex, 20, 1574–1585.
Fahrenfort, J. J., Scholte, H. S., & Lamme, V. A. (2007). Masking disrupts reentrant processing in human visual cortex. Journal of Cognitive Neuroscience, 19, 1488–1497.
Fahrenfort, J. J., Scholte, H. S., & Lamme, V. A. (2008). The spatiotemporal profile of cortical processing leading up to visual perception. Journal of Vision, 8(1, Art. 12), 11–12.
Funes, M. J., Lupiáñez, J., & Milliken, B. (2007). Separate mechanisms recruited by exogenous and endogenous spatial cues: Evidence from a spatial stroop paradigm. Journal of Experimental Psychology: Human Perception and Performance, 33, 348–362.
Hopfinger, J. B., Buonocore, M. H., & Mangun, G. R. (2000). The neural mechanisms of top-down attentional control. Nature Neuroscience, 3, 284–291.
James, W. (1890). The principles of psychology. New York: Holt.
Kentridge, R. W., Heywood, C. A., & Weiskrantz, L. (1999). Attention without awareness in blindsight. Proceedings of rhe Royal Society of London, 266, 1805–1811.
Kentridge, R. W., Heywood, C. A., & Weiskrantz, L. (2004). Spatial attention speeds discrimination without awareness in blindsight. Neuropsychologia, 42, 831–835.
Kentridge, R. W., Nijboer, T. C., & Heywood, C. A. (2008). Attended but unseen: Visual attention is not sufficient for visual awareness. Neuropsychologia, 46, 864–869.
Klein, R. M. (2004). On the control of visual orienting. In M. I. Posner (Ed.), Cognitive neuroscience of attention (pp. 29–44). New York: Guilford.
Koch, C., & Tsuchiya, N. (2007). Attention and consciousness: Two distinct brain processes. Trends in Cognitive Sciences, 11, 16–22.
Kusnir, F., Chica, A. B., Mitsumasu, M. A., & Bartolomeo, P. (2011). Phasic auditory alerting improves visual conscious perception. Consciousness and Cognition.
Lamme, V. A. (2003). Why visual attention and awareness are different. Trends in Cognitive Sciences, 7, 12–18.
Lavie, N. (2006). The role of perceptual load in visual awareness. Brain Research, 1080, 91–100.
Liu, T., Abrams, J., & Carrasco, M. (2009). Voluntary attention enhances contrast appearance. Psychological Science, 20, 354–362.
Lupiáñez, J. (2010). Inhibition of return. In A. C. Nobre & J. T. Coull (Eds.), Attention and time (pp. 17–34). Oxford: Oxford University Press.
Mack, A., & Rock, I. (1998). Inattentional blindness. Cambridge, MA: MIT Press.
McCormick, P. A. (1997). Orienting attention without awareness. Journal of Experimental Psychology: Human Perception and Performance, 23, 168–180.
Mele, S., Savazzi, S., Marzi, C. A., & Berlucchi, G. (2008). Reaction time inhibition from subliminal cues: Is it related to inhibition of return? Neuropsychologia, 46, 810–819.
Merikle, P. M., Smilek, D., & Eastwood, J. D. (2001). Perception without awareness: Perspectives from cognitive psychology. Cognition, 79, 115–134.
Merleau-Ponty, M. (1942). La structure du comportement. Paris: Presses Universitaires de France.
Milliken, B., Tipper, S. P., Houghton, G., & Lupiáñez, J. (2000). Attending, ignoring, and repetition: On the relation between negative priming and inhibition of return. Perception & Psychophysics, 62, 1280–1296.
Mort, D. J., Malhotra, P., Mannan, S. K., Rorden, C., Pambakian, A., Kennard, C., et al. (2003). The anatomy of visual neglect. Brain, 120, 1986–1997.
Müller, H. J., & Rabbitt, P. M. (1989). Reflexive and voluntary orienting of visual attention: Time course of activation and resistance to interruption. Journal of Experimental Psychology: Human Perception and Performance, 15, 315–330.
Nobre, A. C. (2001). The attentive homunculus: Now you see it, now you don't. Neuroscience and Biobehavioral Reviews, 25, 477–496.
O'Regan, J. K., & Noë, A. (2001). A sensorimotor account of vision and visual consciousness. The Behavioral and Brain Sciences, 24, 939–973.
Posner, M. I. (1994). Attention: The mechanisms of consciousness. Proceedings of the National Academy of Sciences, 91, 7398–7403.
Posner, M. I., Snyder, C. R. R., & Davidson, B. J. (1980). Attention and the detection of signals. Journal of Experimental Psychology: General, 109, 160–174.
Prinzmetal, W., McCool, C., & Park, S. (2005). Attention: Reaction time and accuracy reveal different mechanisms. Journal of Experimental Psychology: General, 134, 73–92.
Rolke, B., & Hofmann, P. (2007). Temporal uncertainty degrades perceptual processing. Psychonomic Bulletin & Review, 14, 522–526.
Schneider, W., Eschman, A., & Zuccolotto, A. (2002). E-Prime user’s guide. Pittsburg: Psychology Software Tools.
Sergent, C., Baillet, S., & Dehaene, S. (2005). Timing of the brain events underlying access to consciousness during the attentional blink. Nature Neuroscience, 8, 1391–1400.
Snodgrass, J. G., & Corwin, J. (1988). Pragmatics of measuring recognition memory: Applications to dementia and amnesia. Journal of Experimental Psychology: General, 117, 34–50.
Snodgrass, M., Kalaida, N., & Winer, E. S. (2009). Access is mainly a second-order process: SDT models whether phenomenally (first-order) conscious states are accessed by reflectively (second-order) conscious processes. Consciousness & Cognition, 18, 561–564. discussion 565–567.
Tipples, J. (2002). Eye gaze is not unique: Automatic orienting in response to uninformative arrows. Psychonomic Bulletin & Review, 9, 314–318.
Wilimzig, C., Tsuchiya, N., Fahle, M., Einhauser, W., & Koch, C. (2008). Spatial attention increases performance but not subjective confidence in a discrimination task. Journal of Vision, 8(5, Art. 7), 1–10.
Woodman, G. F., & Luck, S. J. (2003). Dissociations among attention, perception, and awareness during object-substitution masking. Psychological Science, 14, 605–611.
Wyart, V., & Tallon-Baudry, C. (2008). Neural dissociation between visual awareness and spatial attention. The Journal of Neuroscience, 28, 2667–2679.
Acknowledgments
This research was supported by postdoctoral grants from the Neuropôle de Recherche Francilen (NeRF) and Marie Curie Intra-European Program (FP7) to A.B.C., from the Spanish Ministry of Science (research project PSI2008-03595PSIC) to A.B.C. and J.L., and from the Université Pierre et Marie Curie, Paris 6 (Bonus Qualité Recherche) to P.B. The contribution of A.V.-C. was supported by US NINDS R21NSS062317 and FP6 and ANR project eraNET-NEURON BEYONDVIS. L.C. was supported by a predoctoral fellowship from the École des Neurosciences de Paris (ENP, Paris School of Neuroscience).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Chica, A.B., Lasaponara, S., Chanes, L. et al. Spatial attention and conscious perception: the role of endogenous and exogenous orienting. Atten Percept Psychophys 73, 1065–1081 (2011). https://doi.org/10.3758/s13414-010-0082-6
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-010-0082-6